

Regulation of aggregate size and pattern by adenosine and caffeine in cellular slime molds
- PMID: 22269093
- PMCID: PMC3341216
- DOI: 10.1186/1471-213X-12-5
Regulation of aggregate size and pattern by adenosine and caffeine in cellular slime molds
Abstract
Background: Multicellularity in cellular slime molds is achieved by aggregation of several hundreds to thousands of cells. In the model slime mold Dictyostelium discoideum, adenosine is known to increase the aggregate size and its antagonist caffeine reduces the aggregate size. However, it is not clear if the actions of adenosine and caffeine are evolutionarily conserved among other slime molds known to use structurally unrelated chemoattractants. We have examined how the known factors affecting aggregate size are modulated by adenosine and caffeine.
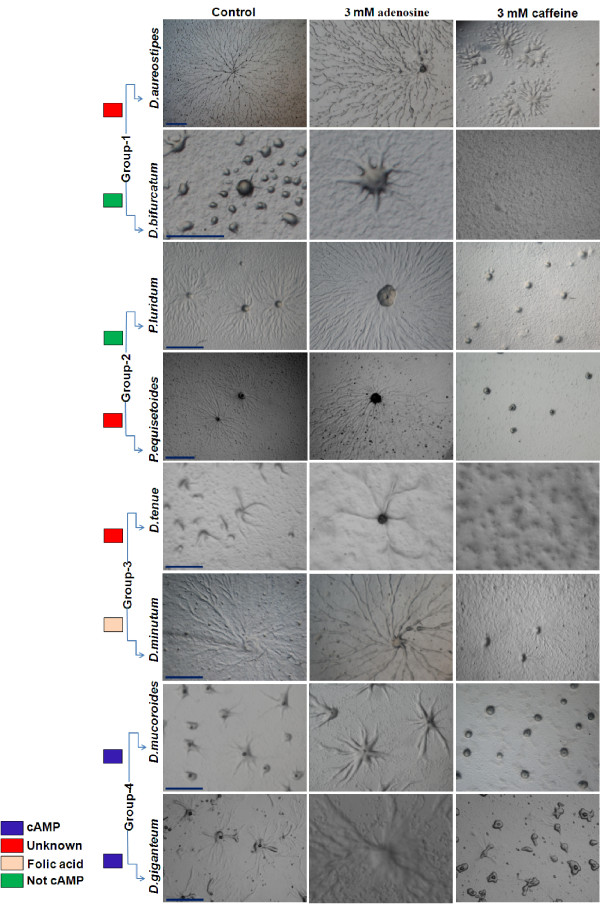
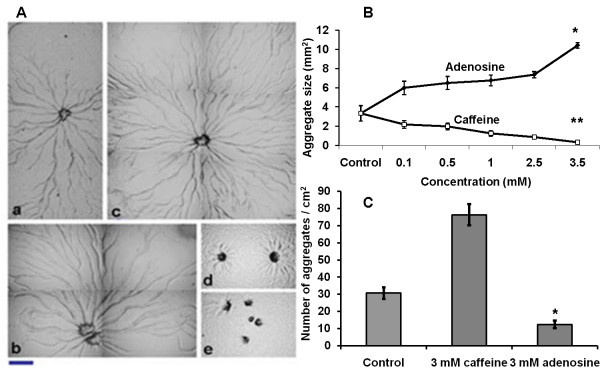
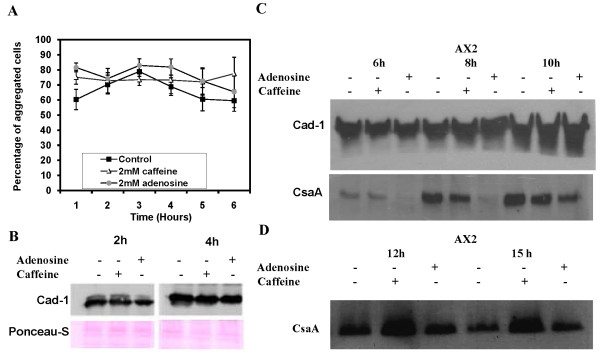
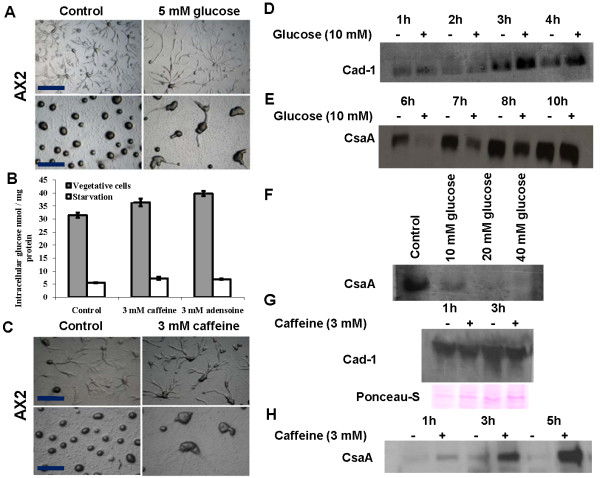
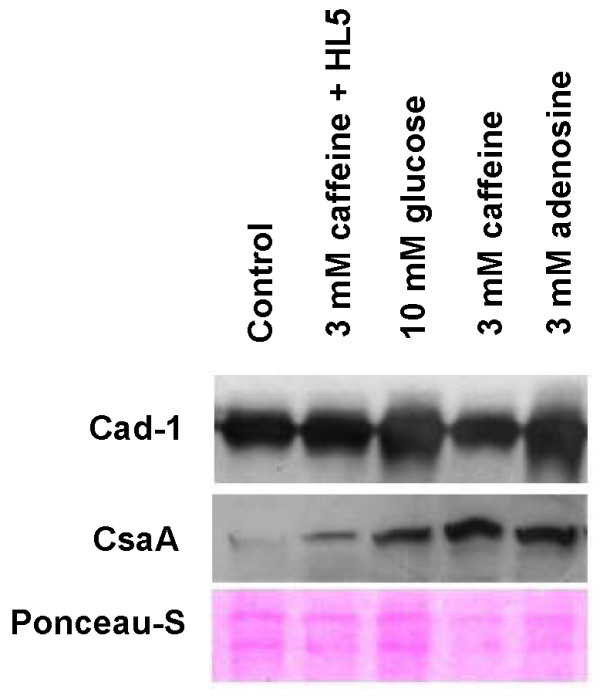
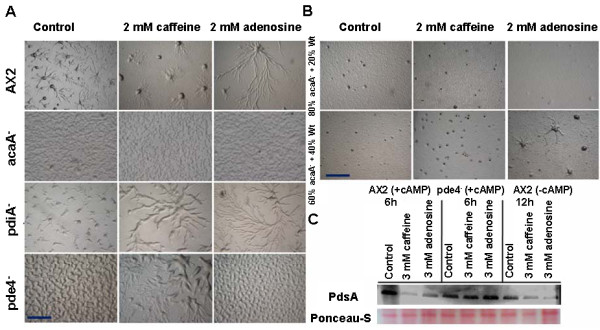
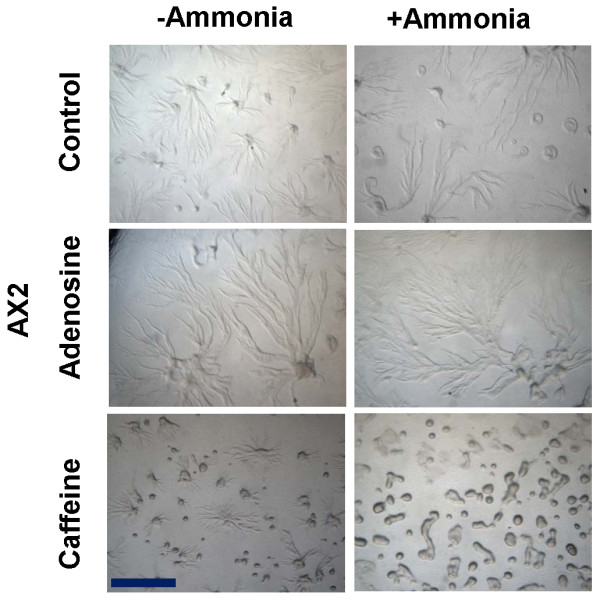
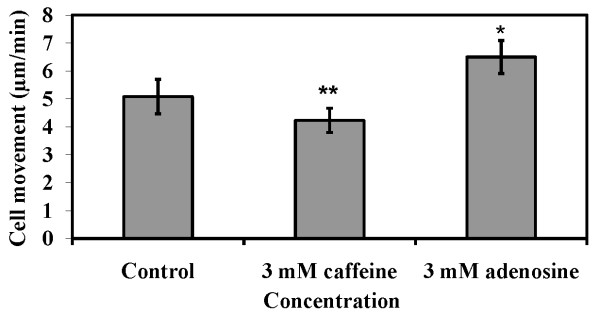
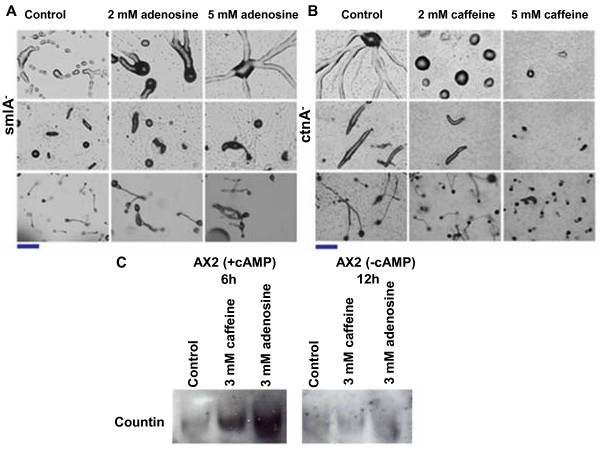
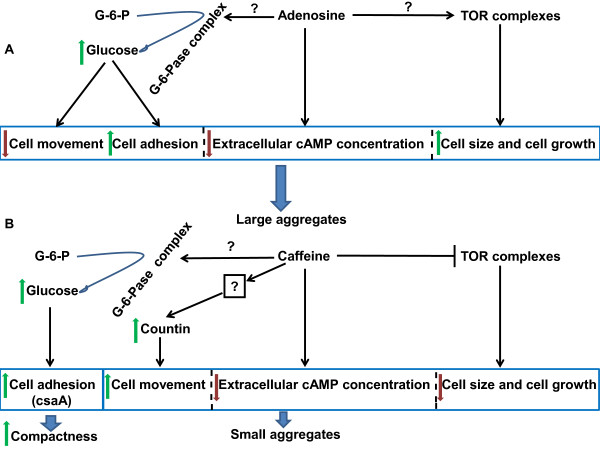
Result: Adenosine and caffeine induced the formation of large and small aggregates respectively, in evolutionarily distinct slime molds known to use diverse chemoattractants for their aggregation. Due to its genetic tractability, we chose D. discoideum to further investigate the factors affecting aggregate size. The changes in aggregate size are caused by the effect of the compounds on several parameters such as cell number and size, cell-cell adhesion, cAMP signal relay and cell counting mechanisms. While some of the effects of these two compounds are opposite to each other, interestingly, both compounds increase the intracellular glucose level and strengthen cell-cell adhesion. These compounds also inhibit the synthesis of cAMP phosphodiesterase (PdsA), weakening the relay of extracellular cAMP signal. Adenosine as well as caffeine rescue mutants impaired in stream formation (pde4- and pdiA-) and colony size (smlA- and ctnA-) and restore their parental aggregate size.
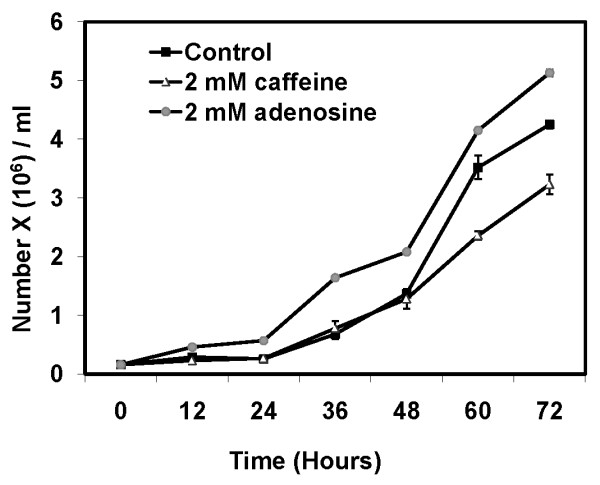
Conclusion: Adenosine increased the cell division timings thereby making large number of cells available for aggregation and also it marginally increased the cell size contributing to large aggregate size. Reduced cell division rates and decreased cell size in the presence of caffeine makes the aggregates smaller than controls. Both the compounds altered the speed of the chemotactic amoebae causing a variation in aggregate size. Our data strongly suggests that cytosolic glucose and extracellular cAMP levels are the other major determinants regulating aggregate size and pattern. Importantly, the aggregation process is conserved among different lineages of cellular slime molds despite using unrelated signalling molecules for aggregation.
Figures
References
-
- Bonner JT. The cellular slime molds. Second. Princeton University Press; 1967.
-
- De Wit RJ, Konijn TM. Identification of the acrasin of Dictyostelium minutum as a derivative of folic acid. Cell Differ. 1983;12:105–210. - PubMed
-
- Konijn TM, Barkley DS, Chang YY, Bonner JT. Cyclic AMP: a naturally occurring acrasin in the cellular slime molds. Am Nat. 1968;102:225–233. doi: 10.1086/282539. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
