

Berardinelli-seip congenital lipodystrophy 2/seipin is a cell-autonomous regulator of lipolysis essential for adipocyte differentiation
- PMID: 22269949
- PMCID: PMC3295006
- DOI: 10.1128/MCB.06465-11
Berardinelli-seip congenital lipodystrophy 2/seipin is a cell-autonomous regulator of lipolysis essential for adipocyte differentiation
Abstract
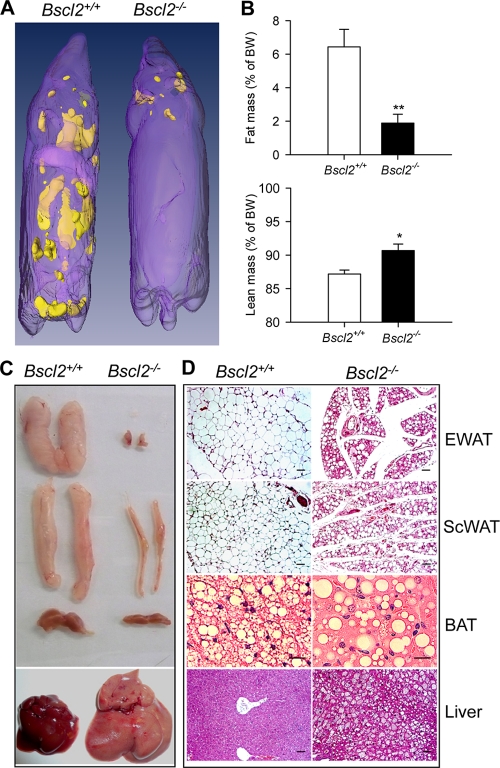
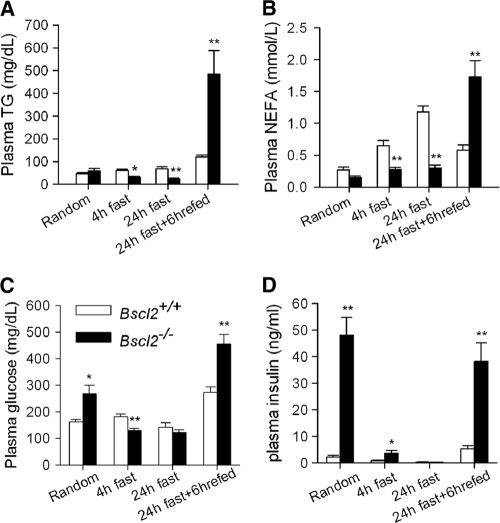
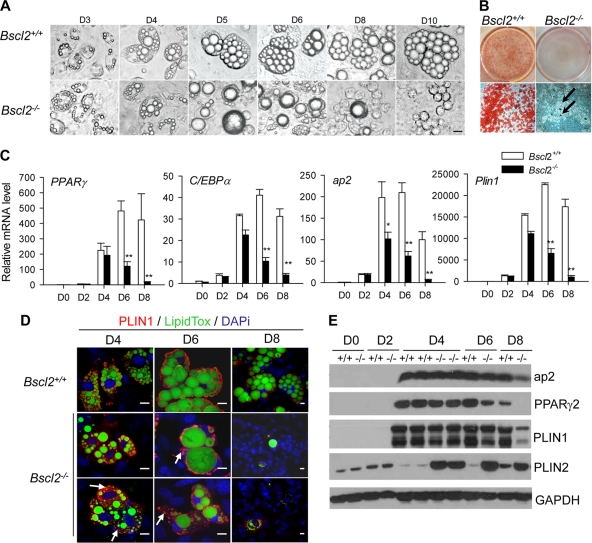
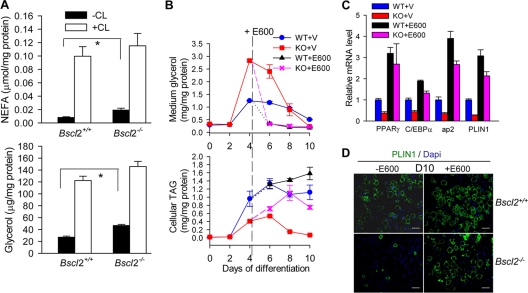
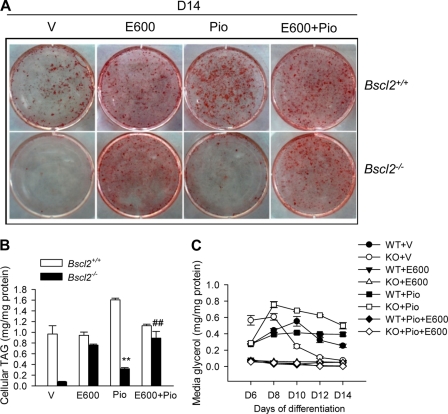
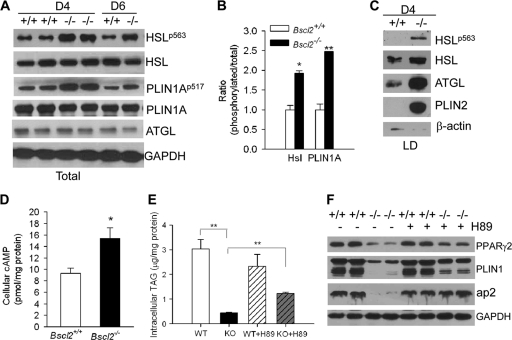
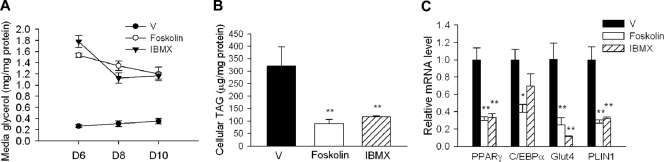
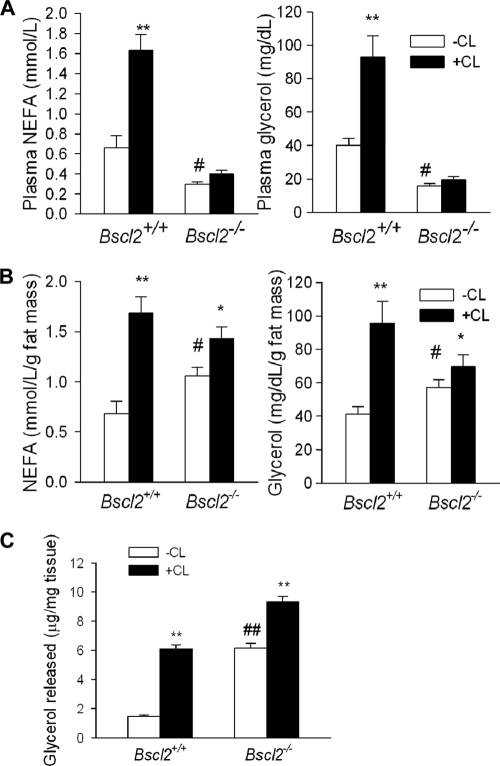
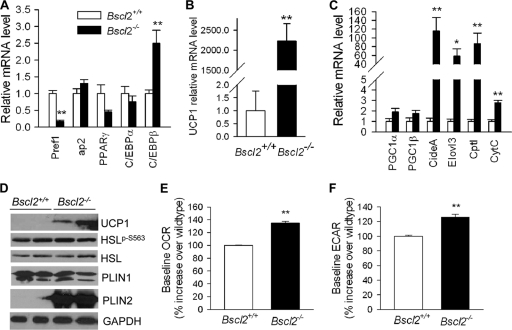
Mutations in BSCL2 underlie human congenital generalized lipodystrophy. We inactivated Bscl2 in mice to examine the mechanisms whereby absence of Bscl2 leads to adipose tissue loss and metabolic disorders. Bscl2(-/-) mice develop severe lipodystrophy of white adipose tissue (WAT), dyslipidemia, insulin resistance, and hepatic steatosis. In vitro differentiation of both Bscl2(-/-) murine embryonic fibroblasts (MEFs) and stromal vascular cells (SVCs) reveals normal early-phase adipocyte differentiation but a striking failure in terminal differentiation due to unbridled cyclic AMP (cAMP)-dependent protein kinase A (PKA)-activated lipolysis, which leads to loss of lipid droplets and silencing of the expression of adipose tissue-specific transcription factors. Importantly, such defects in differentiation can be largely rescued by inhibitors of lipolysis but not by a gamma peroxisome proliferator-activated receptor (PPARγ) agonist. The residual epididymal WAT (EWAT) in Bscl2(-/-) mice displays enhanced lipolysis. It also assumes a "brown-like" phenotype with marked upregulation of UCP1 and other brown adipose tissue-specific markers. Together with decreased Pref1 but increased C/EBPβ levels, these changes highlight a possible increase in cAMP signaling that impairs terminal adipocyte differentiation in the EWAT of Bscl2(-/-) mice. Our study underscores the fundamental role of regulated cAMP/PKA-mediated lipolysis in adipose differentiation and identifies Bscl2 as a novel cell-autonomous determinant of activated lipolysis essential for terminal adipocyte differentiation.
Figures
References
-
- Agarwal AK, Barnes RI, Garg A. 2004. Genetic basis of congenital generalized lipodystrophy. Int. J. Obes. Relat. Metab. Disord. 28:336–339 - PubMed
-
- Agarwal AK, Garg A. 2003. Congenital generalized lipodystrophy: significance of triglyceride biosynthetic pathways. Trends Endocrinol. Metab. 14:214–221 - PubMed
-
- Agarwal AK, et al. 2003. Phenotypic and genetic heterogeneity in congenital generalized lipodystrophy. J. Clin. Endocrinol. Metab. 88:4840–4847 - PubMed
-
- Berardinelli W. 1954. An undiagnosed endocrinometabolic syndrome: report of 2 cases. J. Clin. Endocrinol. Metab. 14:193–204 - PubMed
-
- Bligh EG, Dyer WJ. 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37:911–917 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases