

Proteomic analysis of human hepatoma cells expressing methionine adenosyltransferase I/III: Characterization of DDX3X as a target of S-adenosylmethionine
- PMID: 22270009
- PMCID: PMC4456018
- DOI: 10.1016/j.jprot.2012.01.009
Proteomic analysis of human hepatoma cells expressing methionine adenosyltransferase I/III: Characterization of DDX3X as a target of S-adenosylmethionine
Abstract
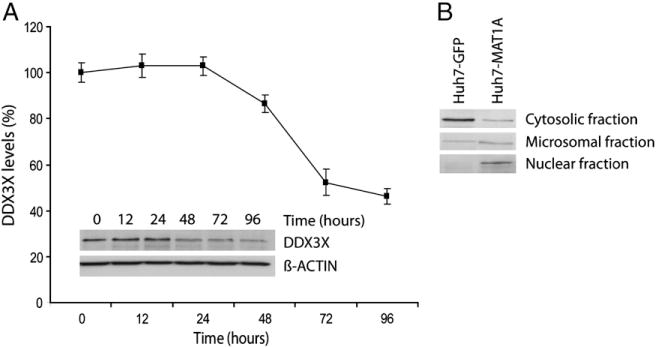
Methionine adenosyltransferase I/III (MATI/III) synthesizes S-adenosylmethionine (SAM) in quiescent hepatocytes. Its activity is compromised in most liver diseases including liver cancer. Since SAM is a driver of hepatocytes fate we have studied the effect of re-expressing MAT1A in hepatoma Huh7 cells using proteomics. MAT1A expression leads to SAM levels close to those found in quiescent hepatocytes and induced apoptosis. Normalization of intracellular SAM induced alteration of 128 proteins identified by 2D-DIGE and gel-free methods, accounting for deregulation of central cellular functions including apoptosis, cell proliferation and survival. Human Dead-box protein 3 (DDX3X), a RNA helicase regulating RNA splicing, export, transcription and translation was down-regulated upon MAT1A expression. Our data support the regulation of DDX3X levels by SAM in a concentration and time dependent manner. Consistently, DDX3X arises as a primary target of SAM and a principal intermediate of its antitumoral effect. Based on the parallelism between SAM and DDX3X along the progression of liver disorders, and the results reported here, it is tempting to suggest that reduced SAM in the liver may lead to DDX3X up-regulation contributing to the pathogenic process and that replenishment of SAM might prove to have beneficial effects, at least in part by reducing DDX3X levels. This article is part of a Special Issue entitled: Proteomics: The clinical link.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures




Similar articles
-
Alterations of Methionine Metabolism as Potential Targets for the Prevention and Therapy of Hepatocellular Carcinoma.Medicina (Kaunas). 2019 Jun 21;55(6):296. doi: 10.3390/medicina55060296. Medicina (Kaunas). 2019. PMID: 31234428 Free PMC article. Review.
-
Pleiotropic effects of methionine adenosyltransferases deregulation as determinants of liver cancer progression and prognosis.J Hepatol. 2013 Oct;59(4):830-41. doi: 10.1016/j.jhep.2013.04.031. Epub 2013 May 7. J Hepatol. 2013. PMID: 23665184 Review.
-
Methionine adenosyltransferase II beta subunit gene expression provides a proliferative advantage in human hepatoma.Gastroenterology. 2003 Apr;124(4):940-8. doi: 10.1053/gast.2003.50151. Gastroenterology. 2003. PMID: 12671891
-
Role of transcriptional and posttranscriptional regulation of methionine adenosyltransferases in liver cancer progression.Hepatology. 2012 Jul;56(1):165-75. doi: 10.1002/hep.25643. Epub 2012 Jun 5. Hepatology. 2012. PMID: 22318685
-
S-adenosyl-L-methionine modifies antioxidant-enzymes, glutathione-biosynthesis and methionine adenosyltransferases-1/2 in hepatitis C virus-expressing cells.World J Gastroenterol. 2016 Apr 14;22(14):3746-57. doi: 10.3748/wjg.v22.i14.3746. World J Gastroenterol. 2016. PMID: 27076759 Free PMC article.
Cited by
-
DDX3X Biomarker Correlates with Poor Survival in Human Gliomas.Int J Mol Sci. 2015 Jul 9;16(7):15578-91. doi: 10.3390/ijms160715578. Int J Mol Sci. 2015. PMID: 26184164 Free PMC article.
-
The influence of neural cell adhesion molecule isoform 140 on the metastasis of thyroid carcinoma.Clin Exp Metastasis. 2013 Mar;30(3):299-307. doi: 10.1007/s10585-012-9537-6. Epub 2012 Sep 27. Clin Exp Metastasis. 2013. PMID: 23015367
-
Conserved metabolite regulation of stress granule assembly via AdoMet.J Cell Biol. 2020 Aug 3;219(8):e201904141. doi: 10.1083/jcb.201904141. J Cell Biol. 2020. PMID: 32609300 Free PMC article.
-
Alterations of Methionine Metabolism as Potential Targets for the Prevention and Therapy of Hepatocellular Carcinoma.Medicina (Kaunas). 2019 Jun 21;55(6):296. doi: 10.3390/medicina55060296. Medicina (Kaunas). 2019. PMID: 31234428 Free PMC article. Review.
-
S-sulfhydration: a cysteine posttranslational modification in plant systems.Plant Physiol. 2015 May;168(1):334-42. doi: 10.1104/pp.15.00009. Epub 2015 Mar 25. Plant Physiol. 2015. PMID: 25810097 Free PMC article.
References
-
- El-Serag HB, Rudolph KL. Hepatocellular carcinoma: epidemiology and molecular carcinogenesis. Gastroenterology. 2007;132:2557–76. - PubMed
-
- Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statistics, 2002. CA Cancer J Clin. 2005;55:74–108. - PubMed
-
- Bosch FX, Ribes J, Borras J. Epidemiology of primary liver cancer. Semin Liver Dis. 1999;19:271–85. - PubMed
-
- Cantoni GL. S-Adenosylmethionine; a new intermediate formed enzymatically from L-methionine and adenosinetriphosphate. J Biol Chem. 1953;204:403–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
