

Constitutive AP-1 activity and EBV infection induce PD-L1 in Hodgkin lymphomas and posttransplant lymphoproliferative disorders: implications for targeted therapy
- PMID: 22271878
- PMCID: PMC3321508
- DOI: 10.1158/1078-0432.CCR-11-1942
Constitutive AP-1 activity and EBV infection induce PD-L1 in Hodgkin lymphomas and posttransplant lymphoproliferative disorders: implications for targeted therapy
Erratum in
- Clin Cancer Res. 2012 Apr 1;18(7):2117
Abstract
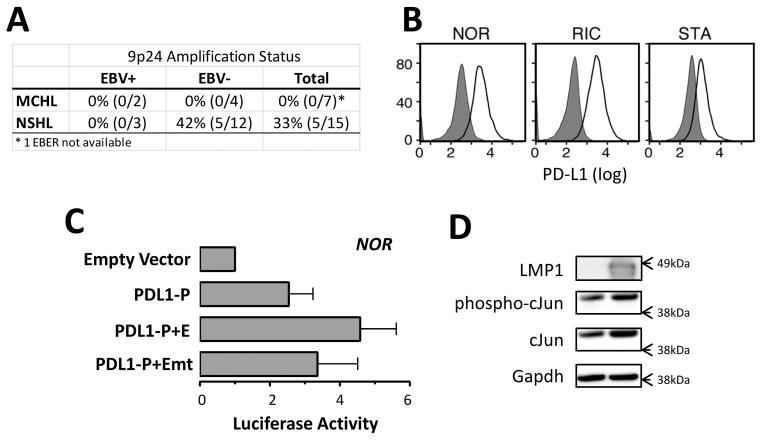
Purpose: Programmed cell death ligand 1 (PD-L1) is a molecule expressed on antigen-presenting cells that engages the PD-1 receptor on T cells and inhibits T-cell receptor signaling. The PD-1 axis can be exploited by tumor cells to dampen host antitumor immune responses and foster tumor cell survival. PD-1 blockade has shown promise in multiple malignancies but should be directed toward patients in whom it will be most effective. In recent studies, we found that the chromosome 9p24.1 amplification increased the gene dosage of PD-L1 and its induction by JAK2 in a subset of patients with classical Hodgkin lymphoma (cHL). However, cHLs with normal 9p24.1 copy numbers also expressed detectable PD-L1, prompting analyses of additional PD-L1 regulatory mechanisms.
Experimental design: Herein, we utilized immunohistochemical, genomic, and functional analyses to define alternative mechanisms of PD-L1 activation in cHL and additional EBV(+) lymphoproliferative disorders.
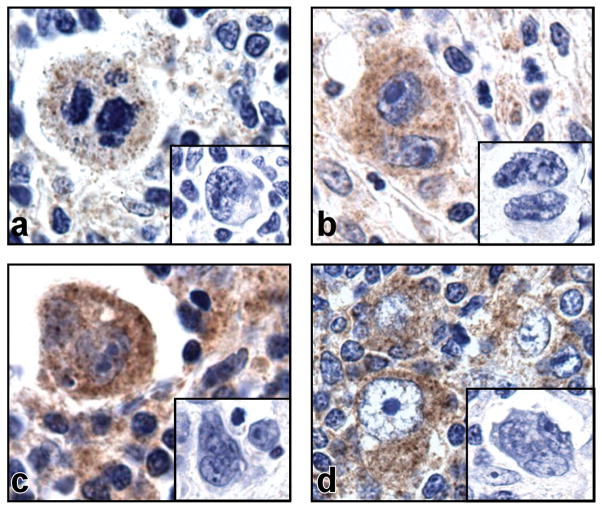
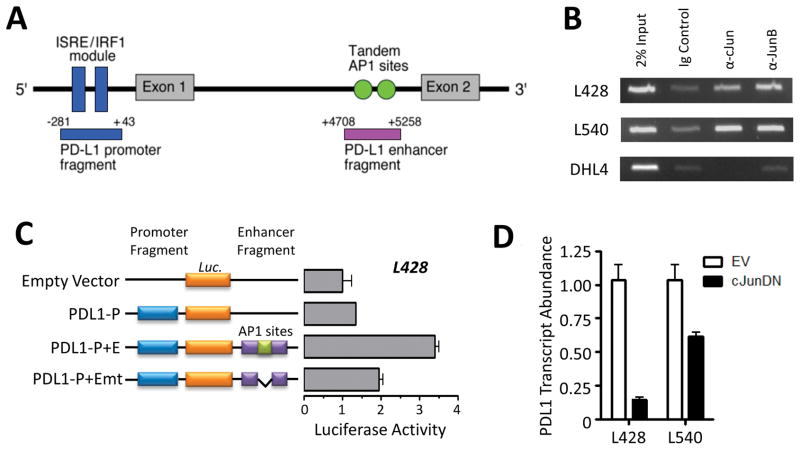
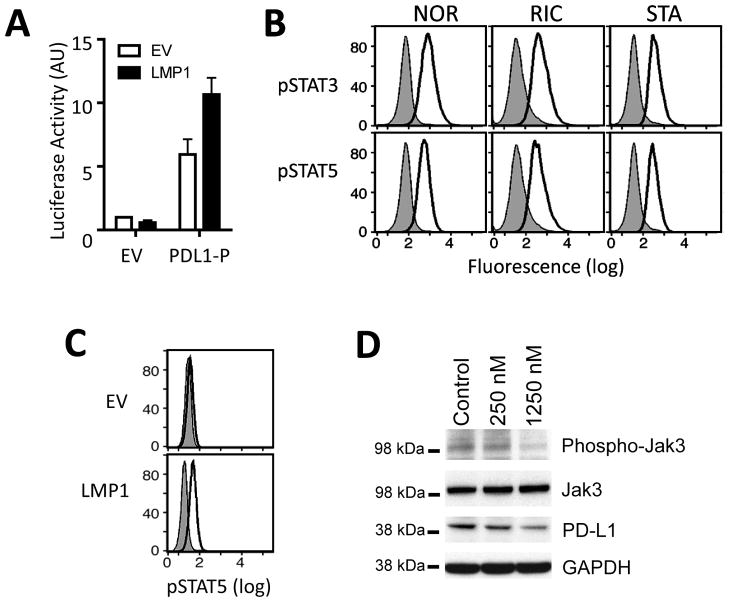
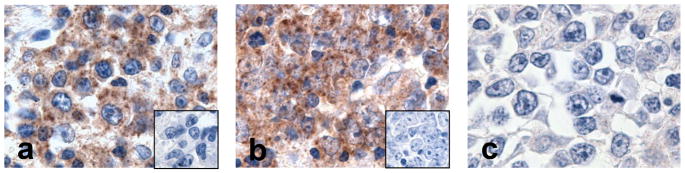
Results: We identified an AP-1-responsive enhancer in the PD-L1 gene. In cHL Reed-Sternberg cells, which exhibit constitutive AP-1 activation, the PD-L1 enhancer binds AP-1 components and increases PD-L1 promoter activity. In addition, we defined Epstein-Barr virus (EBV) infection as an alternative mechanism for PD-L1 induction in cHLs with diploid 9p24.1. PD-L1 was also expressed by EBV-transformed lymphoblastoid cell lines as a result of latent membrane protein 1-mediated, JAK/STAT-dependent promoter and AP-1-associated enhancer activity. In addition, more than 70% of EBV(+) posttransplant lymphoproliferative disorders expressed detectable PD-L1.
Conclusions: AP-1 signaling and EBV infection represent alternative mechanisms of PD-L1 induction and extend the spectrum of tumors in which to consider PD-1 blockade.
Figures
References
-
- Kuppers R. The biology of Hodgkin’s lymphoma. Nat Rev Cancer. 2009;9:15–27. - PubMed
-
- Young L, Rickinson A. Epstein-Barr virus: 40 years on. Nat Rev Cancer. 2004;4(10):757–68. - PubMed
-
- Portis T, Dyck P, Longnecker R. Epstein-Barr Virus (EBV) LMP2A induces alterations in gene transcription similar to those observed in Reed-Sternberg cells of Hodgkin lymphoma. Blood. 2002;102(12):4166–78. - PubMed
-
- Vockerodt M, Morgan S, Kuo M, Wei W, Chukwuma M, Arrand J, et al. The Epstein-Barr virus oncoprotein, latent membrane protein-1 reprograms germinal centre B cells towards a Hodgkin’s Reed-Sternberg-like phenotype. J Pathol. 2008;216(1):83–92. - PubMed
-
- Ouyang J, Juszczynski P, Rodig SJ, Green MR, O’Donnell E, Currie T, et al. Viral induction and targeted inhibition of galectin-1 in EBV+ post-transplant lymphoproliferative disorders. Blood. 117:4315–22. accompanying editorial, Blood 2011; 4317:4165–6, cited in SciBx 4314(4319); doi:101038/scibx2011251 2012011. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
