

Crossing the border: molecular control of motor axon exit
- PMID: 22272090
- PMCID: PMC3257087
- DOI: 10.3390/ijms12128539
Crossing the border: molecular control of motor axon exit
Abstract
Living organisms heavily rely on the function of motor circuits for their survival and for adapting to ever-changing environments. Unique among central nervous system (CNS) neurons, motor neurons (MNs) project their axons out of the CNS. Once in the periphery, motor axons navigate along highly stereotyped trajectories, often at considerable distances from their cell bodies, to innervate appropriate muscle targets. A key decision made by pathfinding motor axons is whether to exit the CNS through dorsal or ventral motor exit points (MEPs). In contrast to the major advances made in understanding the mechanisms that regulate the specification of MN subtypes and the innervation of limb muscles, remarkably little is known about how MN axons project out of the CNS. Nevertheless, a limited number of studies, mainly in Drosophila, have identified transcription factors, and in some cases candidate downstream effector molecules, that are required for motor axons to exit the spinal cord. Notably, specialized neural crest cell derivatives, referred to as Boundary Cap (BC) cells, pre-figure and demarcate MEPs in vertebrates. Surprisingly, however, BC cells are not required for MN axon exit, but rather restrict MN cell bodies from ectopically migrating along their axons out of the CNS. Here, we describe the small set of studies that have addressed motor axon exit in Drosophila and vertebrates, and discuss our fragmentary knowledge of the mechanisms, which guide motor axons out of the CNS.
Keywords: Nkx2.9; Robo; Slit; axon pathfinding; dorsally-exiting motor neuron; motor axon exit; motor exit point; spinal cord; ventrally-exiting motor neuron.
Figures
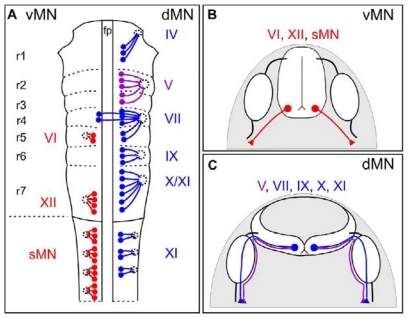
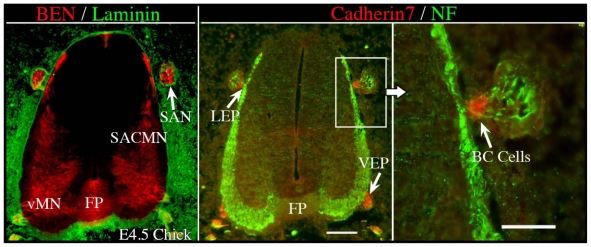
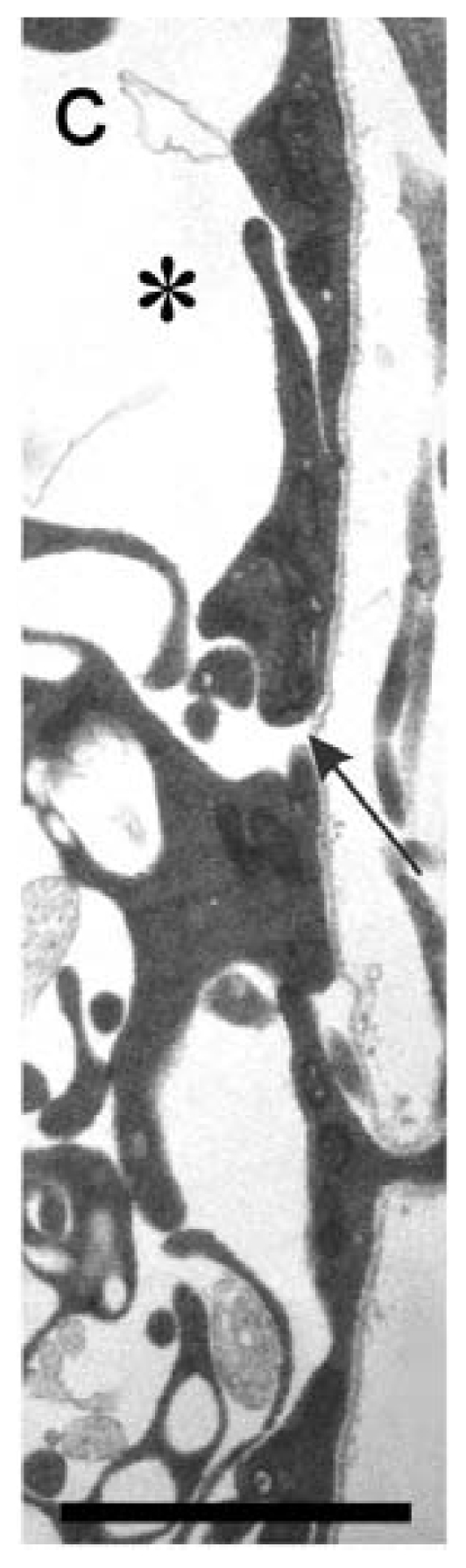
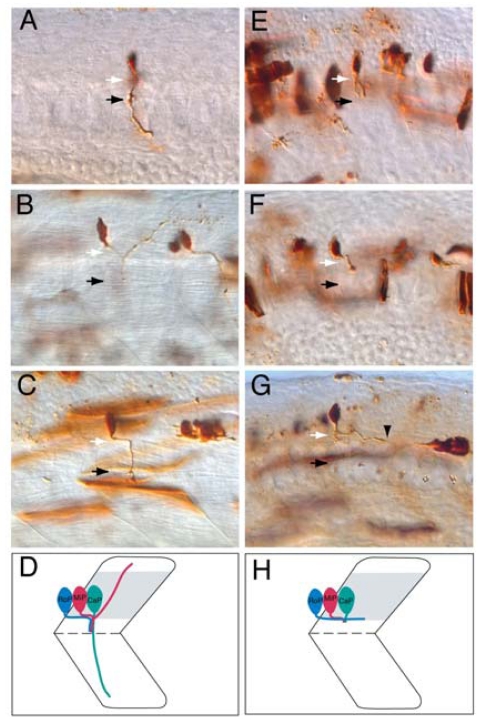
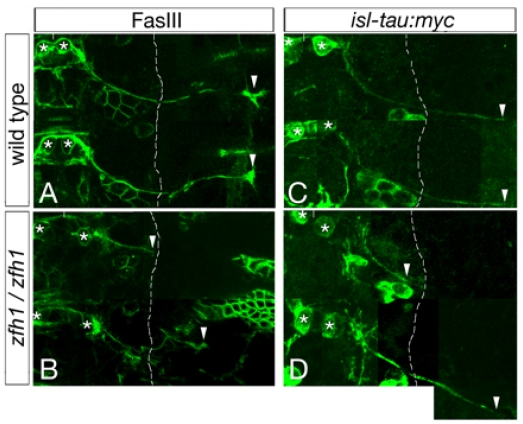
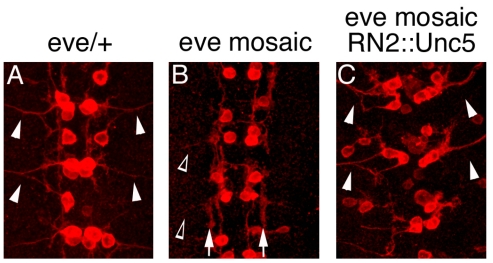
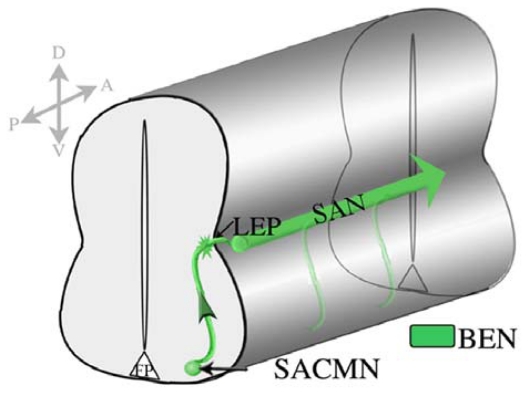
Similar articles
-
Motor axon exit from the mammalian spinal cord is controlled by the homeodomain protein Nkx2.9 via Robo-Slit signaling.Development. 2012 Apr;139(8):1435-46. doi: 10.1242/dev.072256. Epub 2012 Mar 7. Development. 2012. PMID: 22399681 Free PMC article.
-
Boundary cap cells constrain spinal motor neuron somal migration at motor exit points by a semaphorin-plexin mechanism.Neural Dev. 2007 Oct 30;2:21. doi: 10.1186/1749-8104-2-21. Neural Dev. 2007. PMID: 17971221 Free PMC article.
-
Motor axons are guided to exit points in the spinal cord by Slit and Netrin signals.Dev Biol. 2017 Dec 1;432(1):178-191. doi: 10.1016/j.ydbio.2017.09.038. Epub 2017 Oct 3. Dev Biol. 2017. PMID: 28986144 Free PMC article.
-
Motor axon guidance in Drosophila.Semin Cell Dev Biol. 2019 Jan;85:36-47. doi: 10.1016/j.semcdb.2017.11.013. Epub 2017 Nov 24. Semin Cell Dev Biol. 2019. PMID: 29155221 Review.
-
Axon guidance at the midline choice point.Dev Dyn. 2001 Jun;221(2):154-81. doi: 10.1002/dvdy.1143. Dev Dyn. 2001. PMID: 11376484 Review.
Cited by
-
Robo1 and 2 Repellent Receptors Cooperate to Guide Facial Neuron Cell Migration and Axon Projections in the Embryonic Mouse Hindbrain.Neuroscience. 2019 Mar 15;402:116-129. doi: 10.1016/j.neuroscience.2019.01.017. Epub 2019 Jan 24. Neuroscience. 2019. PMID: 30685539 Free PMC article.
-
Motor neuron cell bodies are actively positioned by Slit/Robo repulsion and Netrin/DCC attraction.Dev Biol. 2015 Mar 1;399(1):68-79. doi: 10.1016/j.ydbio.2014.12.014. Epub 2014 Dec 18. Dev Biol. 2015. PMID: 25530182 Free PMC article.
-
Analysis of Expression Pattern and Genetic Deletion of Netrin5 in the Developing Mouse.Front Mol Neurosci. 2016 Jan 26;9:3. doi: 10.3389/fnmol.2016.00003. eCollection 2016. Front Mol Neurosci. 2016. PMID: 26858598 Free PMC article.
-
Lhx3 and Lhx4 suppress Kolmer-Agduhr interneuron characteristics within zebrafish axial motoneurons.Development. 2014 Oct;141(20):3900-9. doi: 10.1242/dev.105718. Epub 2014 Sep 17. Development. 2014. PMID: 25231761 Free PMC article.
-
Motor axon exit from the mammalian spinal cord is controlled by the homeodomain protein Nkx2.9 via Robo-Slit signaling.Development. 2012 Apr;139(8):1435-46. doi: 10.1242/dev.072256. Epub 2012 Mar 7. Development. 2012. PMID: 22399681 Free PMC article.
References
-
- Cordes S.P. Molecular genetics of cranial nerve development in mouse. Nat. Rev. Neurosci. 2001;2:611–623. - PubMed
-
- Schubert W., Kaprielian Z. Identification and characterization of a cell surface marker for embryonic rat spinal accessory motor neurons. J. Comp. Neurol. 2001;439:368–383. - PubMed
-
- Snider W.D., Palavali V. Early axon and dendritic outgrowth of spinal accessory motor neurons studied with DiI in fixed tissues. J. Comp. Neurol. 1990;297:227–238. - PubMed
-
- Sharma K., Sheng H.Z., Lettieri K., Li H., Karavanov A., Potter S., Westphal H., Pfaff S.L. LIM homeodomain factors Lhx3 and Lhx4 assign subtype identities for motor neurons. Cell. 1998;95:817–828. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
