

Sensorimotor function is modulated by the serotonin receptor 1d, a novel marker for gamma motor neurons
- PMID: 22273508
- PMCID: PMC3306528
- DOI: 10.1016/j.mcn.2012.01.003
Sensorimotor function is modulated by the serotonin receptor 1d, a novel marker for gamma motor neurons
Abstract
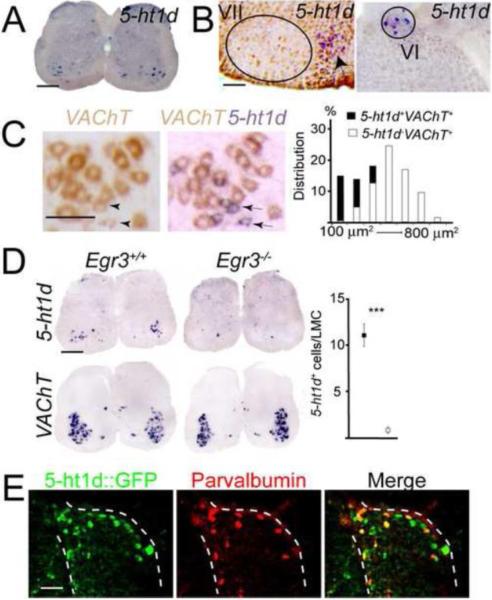
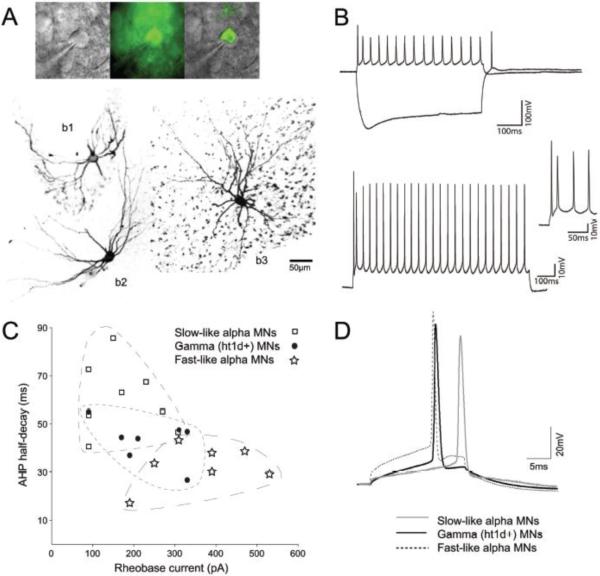
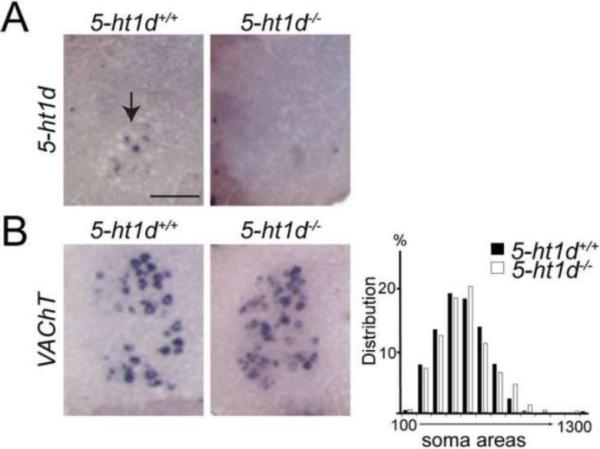
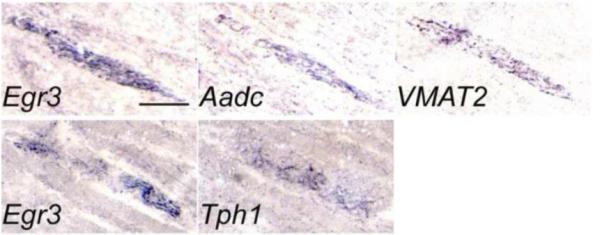
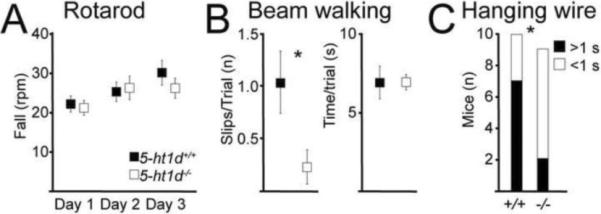
Gamma motor neurons (MNs), the efferent component of the fusimotor system, regulate muscle spindle sensitivity. Muscle spindle sensory feedback is required for proprioception that includes sensing the relative position of neighboring body parts and appropriately adjust the employed strength in a movement. The lack of a single and specific genetic marker has long hampered functional and developmental studies of gamma MNs. Here we show that the serotonin receptor 1d (5-ht1d) is specifically expressed by gamma MNs and proprioceptive sensory neurons. Using mice expressing GFP driven by the 5-ht1d promotor, we performed whole-cell patch-clamp recordings of 5-ht1d::GFP⁺ and 5-ht1d::GFP⁻ motor neurons from young mice. Hierarchal clustering analysis revealed that gamma MNs have distinct electrophysiological properties intermediate to fast-like and slow-like alpha MNs. Moreover, mice lacking 5-ht1d displayed lower monosynaptic reflex amplitudes suggesting a reduced response to sensory stimulation in motor neurons. Interestingly, adult 5-ht1d knockout mice also displayed improved coordination skills on a beam-walking task, implying that reduced activation of MNs by Ia afferents during provoked movement tasks could reduce undesired exaggerated muscle output. In summary, we show that 5-ht1d is a novel marker for gamma MNs and that the 5-ht1d receptor is important for the ability of proprioceptive circuits to receive and relay accurate sensory information in developing and mature spinal cord motor circuits.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Gamma motor neurons express distinct genetic markers at birth and require muscle spindle-derived GDNF for postnatal survival.Neural Dev. 2009 Dec 2;4:42. doi: 10.1186/1749-8104-4-42. Neural Dev. 2009. PMID: 19954518 Free PMC article.
-
Wnt7A identifies embryonic γ-motor neurons and reveals early postnatal dependence of γ-motor neurons on a muscle spindle-derived signal.J Neurosci. 2012 Jun 20;32(25):8725-31. doi: 10.1523/JNEUROSCI.1160-12.2012. J Neurosci. 2012. PMID: 22723712 Free PMC article.
-
Muscle spindle and fusimotor activity in locomotion.J Anat. 2015 Aug;227(2):157-66. doi: 10.1111/joa.12299. Epub 2015 Jun 5. J Anat. 2015. PMID: 26047022 Free PMC article. Review.
-
Topographic studies relating distribution of Ia- and gamma-fibres in spinal cord and position of muscle spindles in cat tibialis anterior muscle.Brain Res. 1985 May 6;333(2):299-304. doi: 10.1016/0006-8993(85)91583-5. Brain Res. 1985. PMID: 3158375
-
On the distribution of information from muscle spindles in the spinal cord; how much does it depend on random factors?J Anat. 2015 Aug;227(2):184-93. doi: 10.1111/joa.12331. J Anat. 2015. PMID: 26179024 Free PMC article. Review.
Cited by
-
ERR2 and ERR3 promote the development of gamma motor neuron functional properties required for proprioceptive movement control.PLoS Biol. 2022 Dec 21;20(12):e3001923. doi: 10.1371/journal.pbio.3001923. eCollection 2022 Dec. PLoS Biol. 2022. PMID: 36542664 Free PMC article.
-
Gamma motor neurons survive and exacerbate alpha motor neuron degeneration in ALS.Proc Natl Acad Sci U S A. 2016 Dec 20;113(51):E8316-E8325. doi: 10.1073/pnas.1605210113. Epub 2016 Dec 7. Proc Natl Acad Sci U S A. 2016. PMID: 27930290 Free PMC article.
-
Role of the trace amine associated receptor 5 (TAAR5) in the sensorimotor functions.Sci Rep. 2021 Nov 29;11(1):23092. doi: 10.1038/s41598-021-02289-w. Sci Rep. 2021. PMID: 34845253 Free PMC article.
-
Functional analysis of human intrafusal fiber innervation by human γ-motoneurons.Sci Rep. 2017 Dec 8;7(1):17202. doi: 10.1038/s41598-017-17382-2. Sci Rep. 2017. PMID: 29222416 Free PMC article.
-
Molecular, Morphological and Electrophysiological Differences between Alpha and Gamma Motoneurons with Special Reference to the Trigeminal Motor Nucleus of Rat.Int J Mol Sci. 2024 May 12;25(10):5266. doi: 10.3390/ijms25105266. Int J Mol Sci. 2024. PMID: 38791305 Free PMC article. Review.
References
-
- Albert J. Baysian Computation with R. 2nd ed. Springer-verlag Gmbh; 2009.
-
- Bonnin A, Torii M, Wang L, Rakic P, Levitt P. Serotonin modulates the response of embryonic thalamocortical axons to netrin-1. Nature neuroscience. 2007;10:588–597. - PubMed
-
- Bryan RN, Trevino DL, Willis WD. Evidence for a common location of alpha and gamma motoneurons. Brain research. 1972;38:193–196. - PubMed
-
- Burke RE, Fyffe RE, Moschovakis AK. Electrotonic architecture of cat gamma motoneurons. Journal of neurophysiology. 1994;72:2302–2316. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
