

SERCA2-controlled Ca²+-dependent keratinocyte adhesion and differentiation is mediated via the sphingolipid pathway: a therapeutic target for Darier's disease
- PMID: 22277942
- PMCID: PMC3305850
- DOI: 10.1038/jid.2011.447
SERCA2-controlled Ca²+-dependent keratinocyte adhesion and differentiation is mediated via the sphingolipid pathway: a therapeutic target for Darier's disease
Abstract
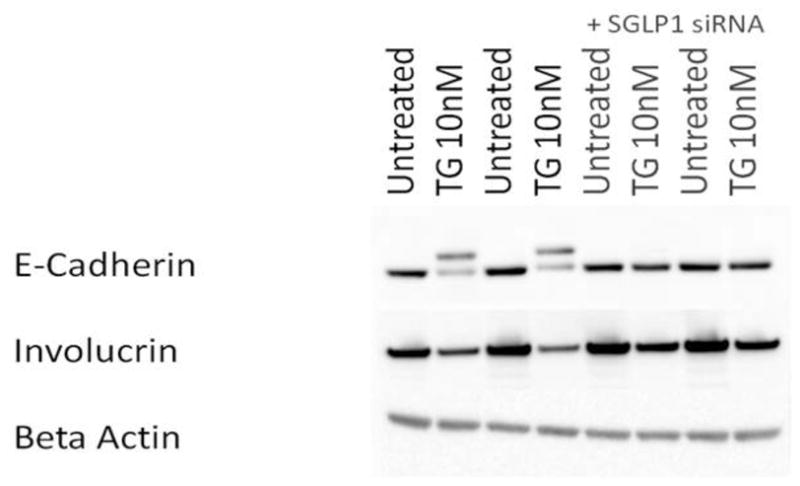
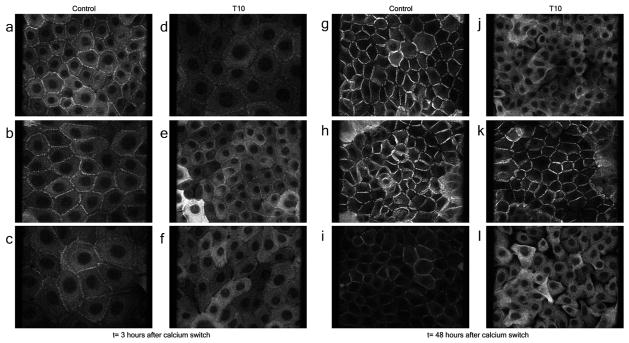
Darier's disease (DD), caused by mutations in the endoplasmic reticulum (ER) Ca(2+) ATPase ATP2A2 (SERCA2b), is a skin disease that exhibits impaired epidermal cell-to-cell adhesion and altered differentiation. Although previous studies have shown that keratinocyte Ca(2+) sequestration and fluxes are controlled by sphingolipid signaling, the role of this signaling pathway in DD previously has not been investigated. We show here that sphingosine levels increase and sphingosine kinase (SPHK1) expression decreases after inactivating SERCA2b with the specific SERCA2 inhibitors thapsigargin (TG) or small interfering RNA to SERCA2b. Conversely, inhibiting sphingosine lyase rescues the defects in keratinocyte differentiation, E-cadherin localization, desmoplakin (DP) translocation, and ER Ca(2+) sequestration seen in TG-treated keratinocytes. Here, we report early evidence that the keratinocyte sphingolipid and Ca(2+) signaling pathways intersect in ATP2A2-controlled ER Ca(2+) sequestration, E-cadherin and DP localization, and Ca(2+)-controlled differentiation, and thus may be important mediators in DD.
Conflict of interest statement
Figures





Similar articles
-
SERCA2 dysfunction in Darier disease causes endoplasmic reticulum stress and impaired cell-to-cell adhesion strength: rescue by Miglustat.J Invest Dermatol. 2014 Jul;134(7):1961-1970. doi: 10.1038/jid.2014.8. Epub 2014 Jan 3. J Invest Dermatol. 2014. PMID: 24390139
-
Impaired trafficking of the desmoplakins in cultured Darier's disease keratinocytes.J Invest Dermatol. 2003 Dec;121(6):1349-55. doi: 10.1046/j.1523-1747.2003.12557.x. J Invest Dermatol. 2003. PMID: 14675181
-
The calcium ATPase SERCA2 regulates desmoplakin dynamics and intercellular adhesive strength through modulation of PKCα signaling.FASEB J. 2011 Mar;25(3):990-1001. doi: 10.1096/fj.10-163261. Epub 2010 Dec 14. FASEB J. 2011. PMID: 21156808 Free PMC article.
-
Markers of squamous cell carcinoma in sarco/endoplasmic reticulum Ca2+ ATPase 2 heterozygote mice keratinocytes.Prog Biophys Mol Biol. 2010 Sep;103(1):81-7. doi: 10.1016/j.pbiomolbio.2009.10.005. Epub 2009 Oct 17. Prog Biophys Mol Biol. 2010. PMID: 19840814 Review.
-
Darier disease : a disease model of impaired calcium homeostasis in the skin.Biochim Biophys Acta. 2011 May;1813(5):1111-7. doi: 10.1016/j.bbamcr.2010.12.006. Epub 2010 Dec 15. Biochim Biophys Acta. 2011. PMID: 21167218 Review.
Cited by
-
Cell cycle controls long-range calcium signaling in the regenerating epidermis.J Cell Biol. 2023 Jul 3;222(7):e202302095. doi: 10.1083/jcb.202302095. Epub 2023 Apr 27. J Cell Biol. 2023. PMID: 37102999 Free PMC article.
-
Genetic pathways in disorders of epidermal differentiation.Trends Genet. 2013 Jan;29(1):31-40. doi: 10.1016/j.tig.2012.10.005. Epub 2012 Nov 8. Trends Genet. 2013. PMID: 23141808 Free PMC article. Review.
-
Genetics of Darier's Disease: New Insights into Pathogenic Mechanisms.Genes (Basel). 2025 May 23;16(6):619. doi: 10.3390/genes16060619. Genes (Basel). 2025. PMID: 40565511 Free PMC article. Review.
-
Pumping the Breaks on Acantholytic Skin Disorders: Targeting Calcium Pumps, Desmosomes, and Downstream Signaling in Darier, Hailey-Hailey, and Grover Disease.J Invest Dermatol. 2025 Mar;145(3):494-508. doi: 10.1016/j.jid.2024.06.1289. Epub 2024 Aug 29. J Invest Dermatol. 2025. PMID: 39207315 Review.
-
Darier disease: first molecular study of a Portuguese family.Heliyon. 2019 Sep 26;5(9):e02520. doi: 10.1016/j.heliyon.2019.e02520. eCollection 2019 Sep. Heliyon. 2019. PMID: 31687605 Free PMC article.
References
-
- Bikle DD, Ng D, Tu CL, et al. Calcium- and vitamin D-regulated keratinocyte differentiation. Mol Cell Endocrinol. 2001;177:161–171. - PubMed
-
- Callewaert G, Parys JB, De Smedt H, et al. Similar Ca(2+)-signaling properties in keratinocytes and in COS-1 cells overexpressing the secretory-pathway Ca(2+)-ATPase SPCA1. Cell Calcium. 2003;34:157–162. - PubMed
-
- Claas RF, ter Braak M, Hegen B, et al. Enhanced Ca2+ storage in sphingosine-1-phosphate lyase-deficient fibroblasts. Cell Signal. 2010;22:476–483. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
