

Calcium binding by synaptotagmin's C2A domain is an essential element of the electrostatic switch that triggers synchronous synaptic transmission
- PMID: 22279210
- PMCID: PMC3567453
- DOI: 10.1523/JNEUROSCI.4652-11.2012
Calcium binding by synaptotagmin's C2A domain is an essential element of the electrostatic switch that triggers synchronous synaptic transmission
Abstract
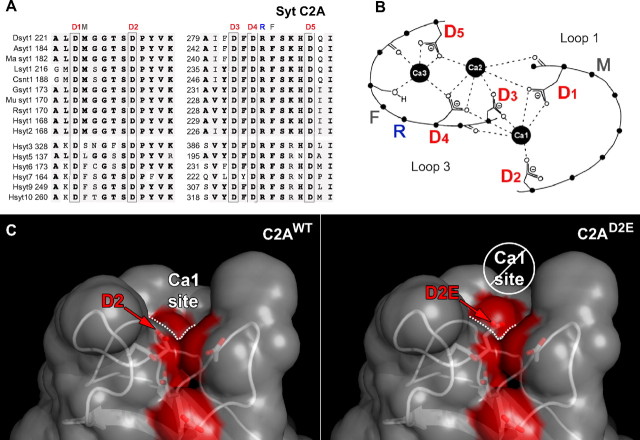
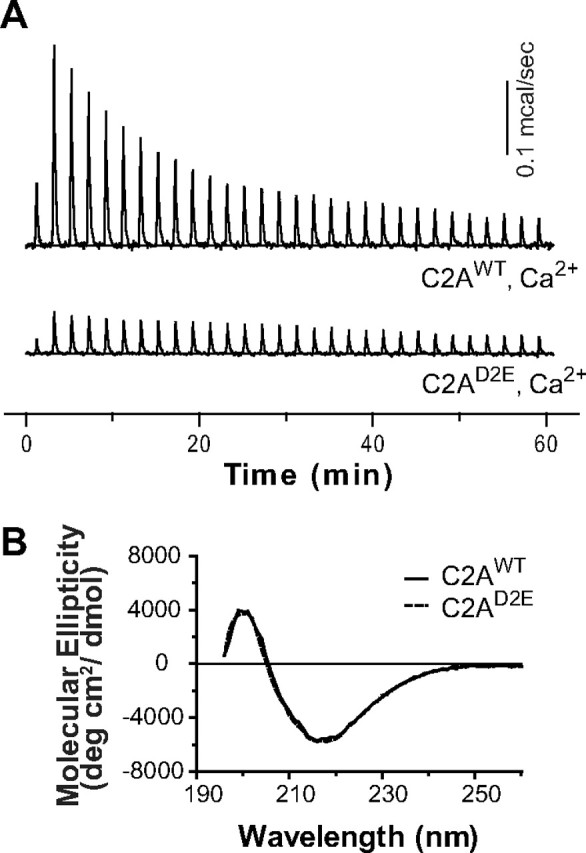
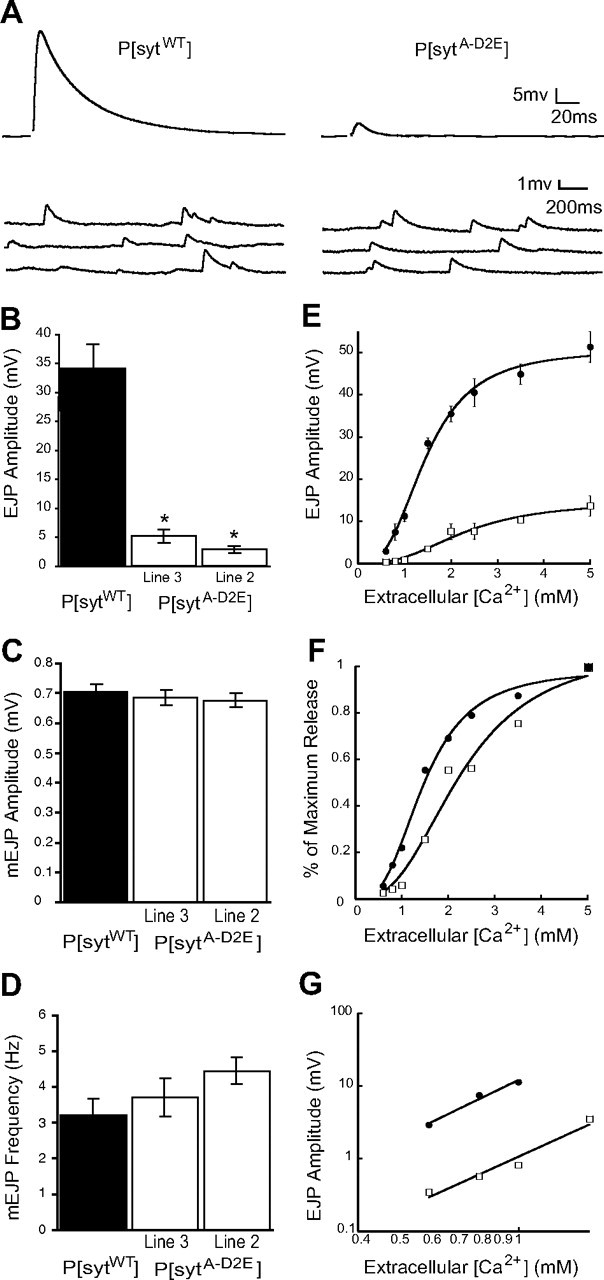
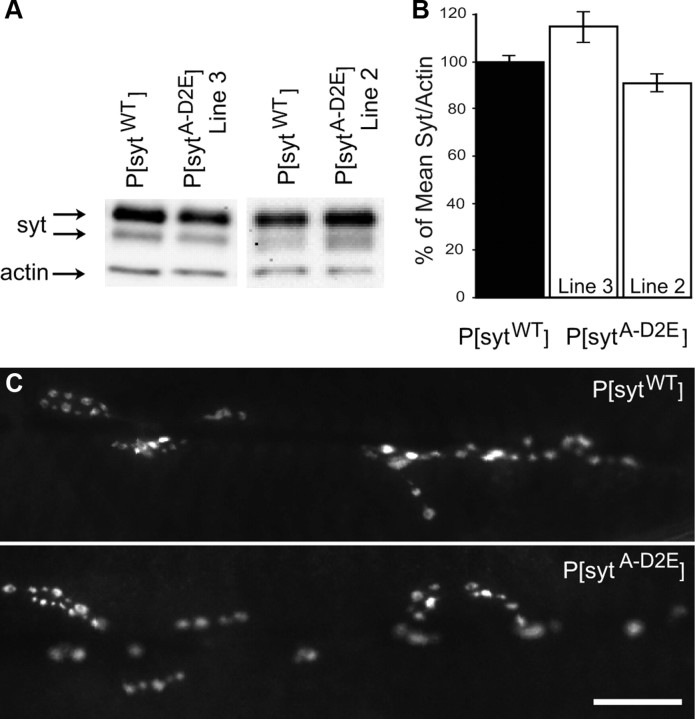
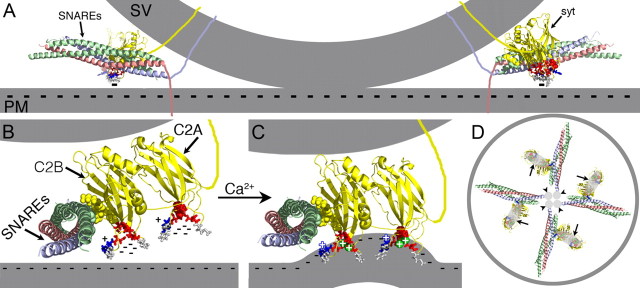
Synaptotagmin is the major calcium sensor for fast synaptic transmission that requires the synchronous fusion of synaptic vesicles. Synaptotagmin contains two calcium-binding domains: C2A and C2B. Mutation of a positively charged residue (R233Q in rat) showed that Ca2+-dependent interactions between the C2A domain and membranes play a role in the electrostatic switch that initiates fusion. Surprisingly, aspartate-to-asparagine mutations in C2A that inhibit Ca2+ binding support efficient synaptic transmission, suggesting that Ca2+ binding by C2A is not required for triggering synchronous fusion. Based on a structural analysis, we generated a novel mutation of a single Ca2+-binding residue in C2A (D229E in Drosophila) that inhibited Ca2+ binding but maintained the negative charge of the pocket. This C2A aspartate-to-glutamate mutation resulted in ∼80% decrease in synchronous transmitter release and a decrease in the apparent Ca2+ affinity of release. Previous aspartate-to-asparagine mutations in C2A partially mimicked Ca2+ binding by decreasing the negative charge of the pocket. We now show that the major function of Ca2+ binding to C2A is to neutralize the negative charge of the pocket, thereby unleashing the fusion-stimulating activity of synaptotagmin. Our results demonstrate that Ca2+ binding by C2A is a critical component of the electrostatic switch that triggers synchronous fusion. Thus, Ca2+ binding by C2B is necessary and sufficient to regulate the precise timing required for coupling vesicle fusion to Ca2+ influx, but Ca2+ binding by both C2 domains is required to flip the electrostatic switch that triggers efficient synchronous synaptic transmission.
Figures
References
-
- Bhalla A, Chicka MC, Tucker WC, Chapman ER. Ca(2+)-synaptotagmin directly regulates t-SNARE function during reconstituted membrane fusion. Nat Struct Mol Biol. 2006;13:323–330. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Brose N, Petrenko AG, Südhof TC, Jahn R. Synaptotagmin: a calcium sensor on the synaptic vesicle surface. Science. 1992;256:1021–1025. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous