

Transgenic expression of intraneuronal Aβ42 but not Aβ40 leads to cellular Aβ lesions, degeneration, and functional impairment without typical Alzheimer's disease pathology
- PMID: 22279212
- PMCID: PMC6796269
- DOI: 10.1523/JNEUROSCI.4586-11.2012
Transgenic expression of intraneuronal Aβ42 but not Aβ40 leads to cellular Aβ lesions, degeneration, and functional impairment without typical Alzheimer's disease pathology
Abstract
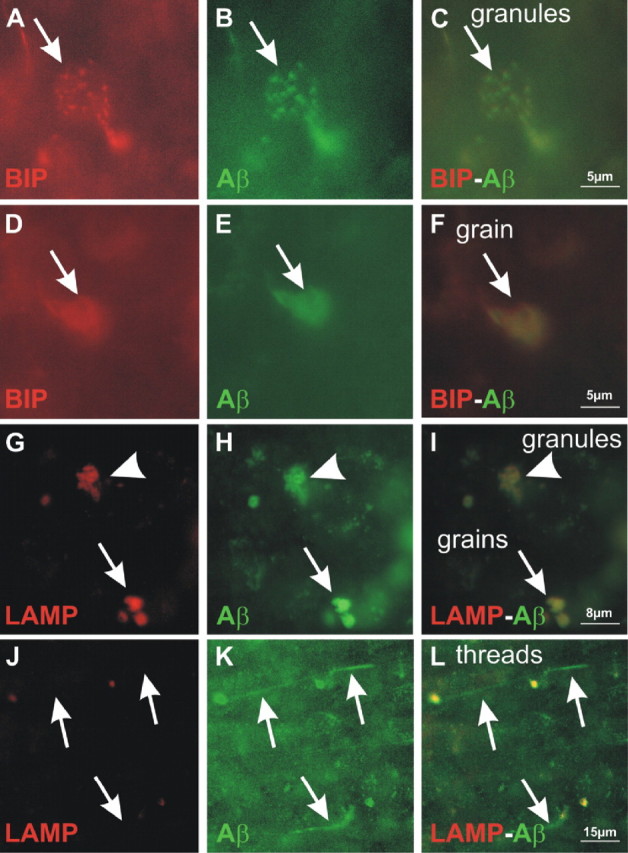
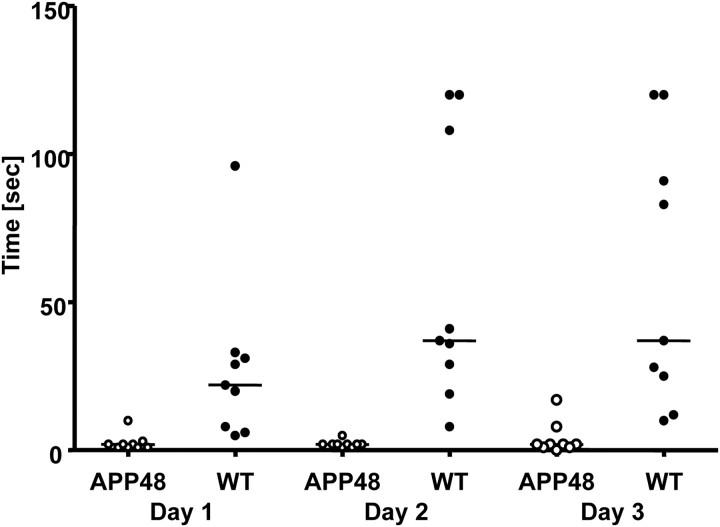
An early role of amyloid-β peptide (Aβ) aggregation in Alzheimer's disease pathogenesis is well established. However, the contribution of intracellular or extracellular forms of Aβ to the neurodegenerative process is a subject of considerable debate. We here describe transgenic mice expressing Aβ1-40 (APP47) and Aβ1-42 (APP48) with a cleaved signal sequence to insert both peptides during synthesis into the endoplasmic reticulum. Although lower in transgene mRNA, APP48 mice reach a higher brain Aβ concentration. The reduced solubility and increased aggregation of Aβ1-42 may impair its degradation. APP48 mice develop intracellular Aβ lesions in dendrites and lysosomes. The hippocampal neuron number is reduced already at young age. The brain weight decreases during aging in conjunction with severe white matter atrophy. The mice show a motor impairment. Only very few Aβ1-40 lesions are found in APP47 mice. Neither APP47 nor APP48 nor the bigenic mice develop extracellular amyloid plaques. While intracellular membrane expression of Aβ1-42 in APP48 mice does not lead to the AD-typical lesions, Aβ aggregates develop within cells accompanied by considerable neurodegeneration.
Figures








References
-
- Abramowski D, Wiederhold KH, Furrer U, Jaton AL, Neuenschwander A, Runser MJ, Danner S, Reichwald J, Ammaturo D, Staab D, Stoeckli M, Rueeger H, Neumann U, Staufenbiel M. Dynamics of A beta turnover and deposition in different beta-amyloid precursor protein transgenic mouse models following gamma-secretase inhibition. J Pharmacol Exp Ther. 2008;327:411–424. - PubMed
-
- Braak H. On the structure of the human archicortex. I. The cornu ammonis. A Golgi and pigmentarchitectonic study. Cell Tissue Res. 1974;152:349–383. - PubMed
-
- Braak H, Braak E. Demonstration of amyloid deposits and neurofibrillary changes in whole brain sections. Brain Pathol. 1991;1:213–216. - PubMed
-
- Calhoun ME, Wiederhold KH, Abramowski D, Phinney AL, Probst A, Sturchler-Pierrat C, Staufenbiel M, Sommer B, Jucker M. Neuron loss in APP transgenic mice. Nature. 1998;395:755–756. - PubMed
-
- Calhoun ME, Burgermeister P, Phinney AL, Stalder M, Tolnay M, Wiederhold KH, Abramowski D, Sturchler-Pierrat C, Sommer B, Staufenbiel M, Jucker M. Neuronal overexpression of mutant amyloid precursor protein results in prominent deposition of cerebrovascular amyloid. Proc Natl Acad Sci U S A. 1999;96:14088–14093. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases