

Subtotal ablation of parietal epithelial cells induces crescent formation
- PMID: 22282596
- PMCID: PMC3312496
- DOI: 10.1681/ASN.2011050449
Subtotal ablation of parietal epithelial cells induces crescent formation
Abstract
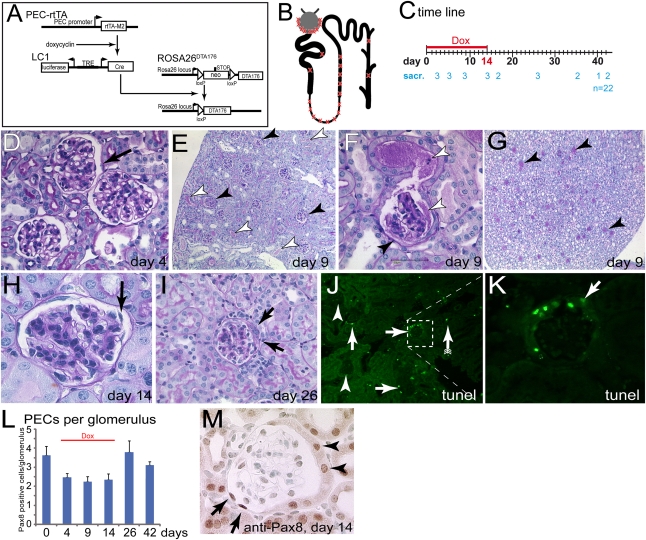
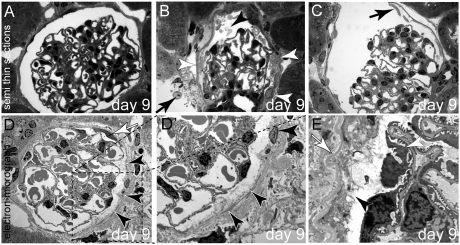
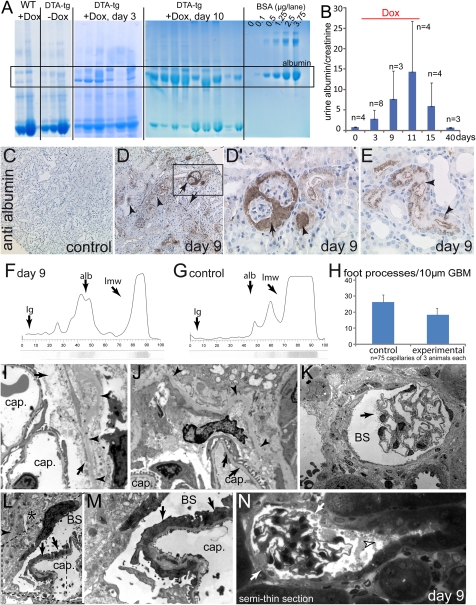
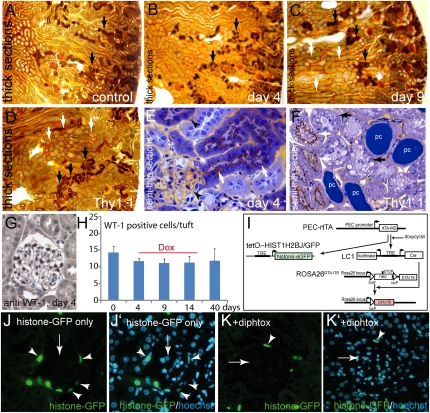
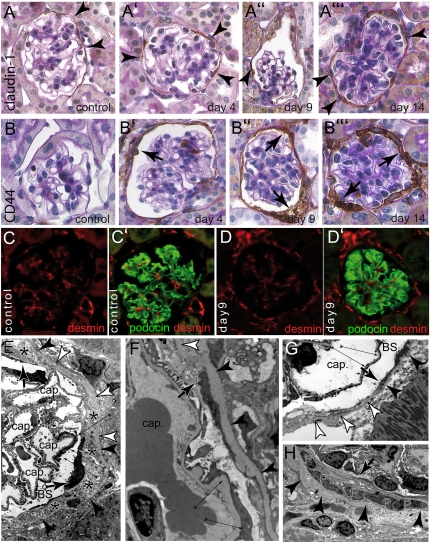
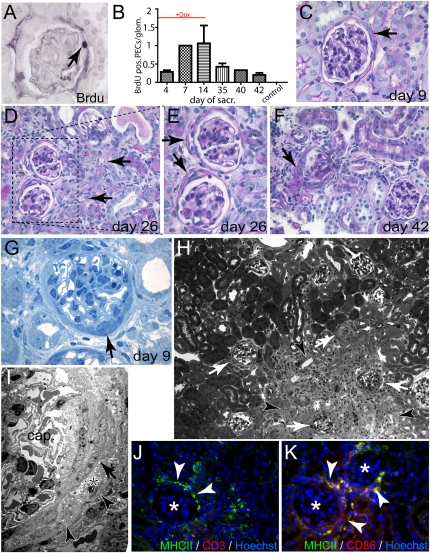
Parietal epithelial cells (PECs) of the renal glomerulus contribute to the formation of both cellular crescents in rapidly progressive GN and sclerotic lesions in FSGS. Subtotal transgenic ablation of podocytes induces FSGS but the effect of specific ablation of PECs is unknown. Here, we established an inducible transgenic mouse to allow subtotal ablation of PECs. Proteinuria developed during doxycycline-induced cellular ablation but fully reversed 26 days after termination of doxycycline administration. The ablation of PECs was focal, with only 30% of glomeruli exhibiting histologic changes; however, the number of PECs was reduced up to 90% within affected glomeruli. Ultrastructural analysis revealed disruption of PEC plasma membranes with cytoplasm shedding into Bowman's space. Podocytes showed focal foot process effacement, which was the most likely cause for transient proteinuria. After >9 days of cellular ablation, the remaining PECs formed cellular extensions to cover the denuded Bowman's capsule and expressed the activation marker CD44 de novo. The induced proliferation of PECs persisted throughout the observation period, resulting in the formation of typical cellular crescents with periglomerular infiltrate, albeit without accompanying proteinuria. In summary, subtotal ablation of PECs leads the remaining PECs to react with cellular activation and proliferation, which ultimately forms cellular crescents.
Figures
References
-
- Moeller MJ, Soofi A, Hartmann I, Le Hir M, Wiggins R, Kriz W, Holzman LB: Podocytes populate cellular crescents in a murine model of inflammatory glomerulonephritis. J Am Soc Nephrol 15: 61–67, 2004 - PubMed
-
- Ding M, Cui S, Li C, Jothy S, Haase V, Steer BM, Marsden PA, Pippin J, Shankland S, Rastaldi MP, Cohen CD, Kretzler M, Quaggin SE: Loss of the tumor suppressor Vhlh leads to upregulation of Cxcr4 and rapidly progressive glomerulonephritis in mice. Nat Med 12: 1081–1087, 2006 - PubMed
-
- Smeets B, Kuppe C, Sicking EM, Fuss A, Jirak P, van Kuppelvelt TH, Endlich KH, Wetzels JF, Grone HJ, Floege J, Moeller MJ: Parietal epithelial cells participate in the formation of sclerotic lesions in focal and segmental glomerulosclerosis (FSGS). J Am Soc Nephrol 22: 1262-1274, 2011 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
