

Postsynaptic complexin controls AMPA receptor exocytosis during LTP
- PMID: 22284181
- PMCID: PMC3269030
- DOI: 10.1016/j.neuron.2011.11.020
Postsynaptic complexin controls AMPA receptor exocytosis during LTP
Abstract
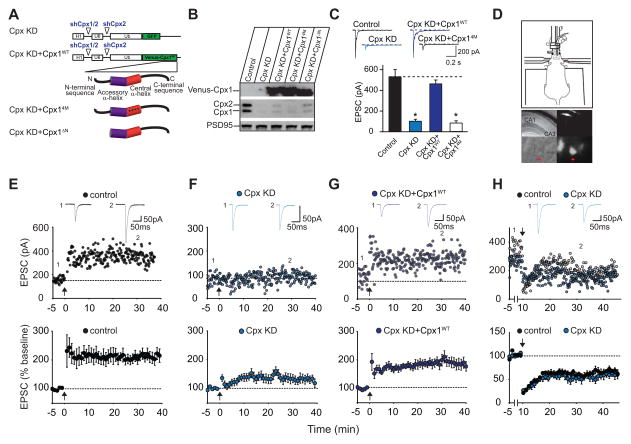
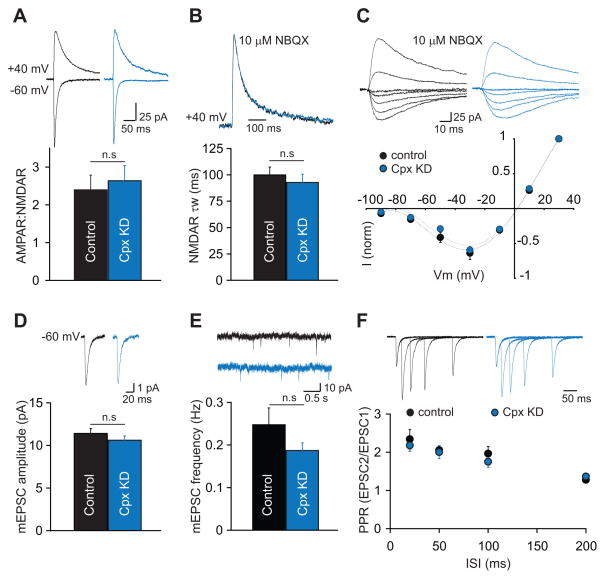
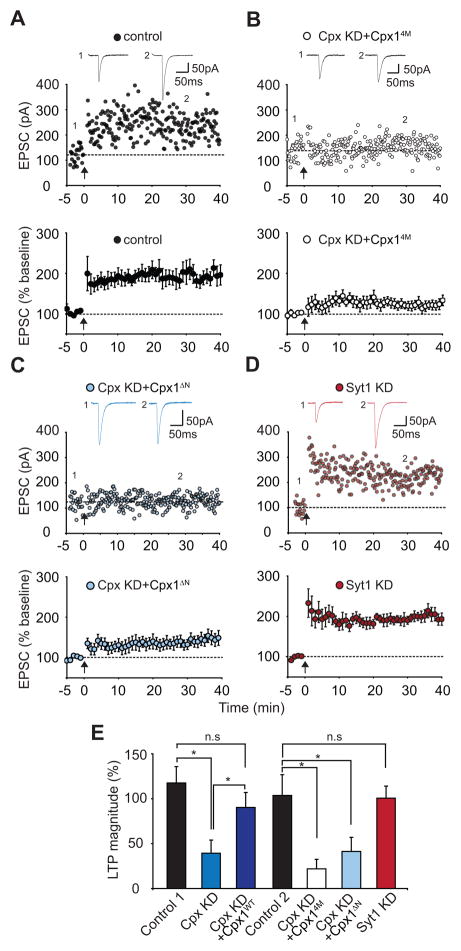
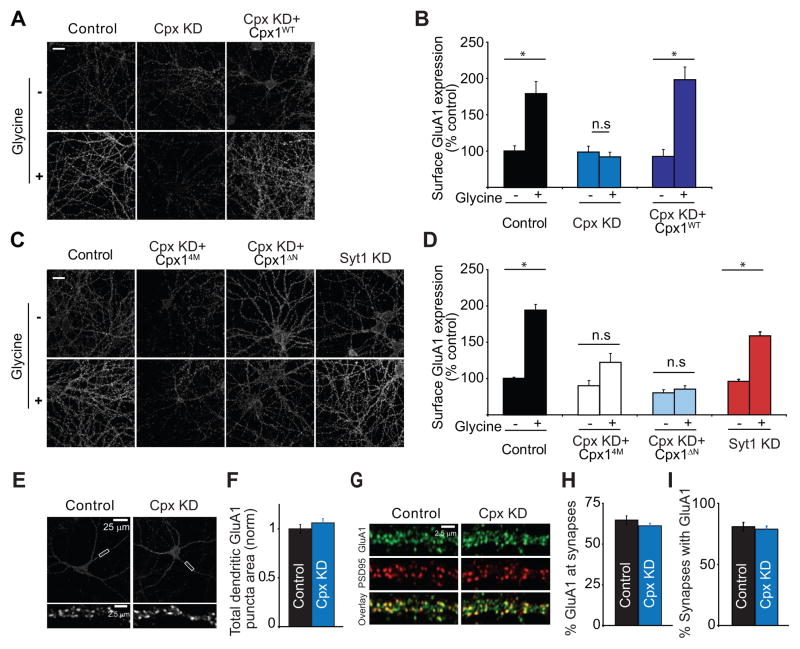
Long-term potentiation (LTP) is a compelling synaptic correlate of learning and memory. LTP induction requires NMDA receptor (NMDAR) activation, which triggers SNARE-dependent exocytosis of AMPA receptors (AMPARs). However, the molecular mechanisms mediating AMPAR exocytosis induced by NMDAR activation remain largely unknown. Here, we show that complexin, a protein that regulates neurotransmitter release via binding to SNARE complexes, is essential for AMPAR exocytosis during LTP but not for the constitutive AMPAR exocytosis that maintains basal synaptic strength. The regulated postsynaptic AMPAR exocytosis during LTP requires binding of complexin to SNARE complexes. In hippocampal neurons, presynaptic complexin acts together with synaptotagmin-1 to mediate neurotransmitter release. However, postsynaptic synaptotagmin-1 is not required for complexin-dependent AMPAR exocytosis during LTP. These results suggest a complexin-dependent molecular mechanism for regulating AMPAR delivery to synapses, a mechanism that is surprisingly similar to presynaptic exocytosis but controlled by regulators other than synaptotagmin-1.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Synaptic plasticity: Adding a piece to the LTP jigsaw.Nat Rev Neurosci. 2012 Feb 15;13(3):150-1. doi: 10.1038/nrn3207. Nat Rev Neurosci. 2012. PMID: 22334211 No abstract available.
References
-
- Adesnik H, Nicoll RA, England PM. Photoinactivation of native AMPA receptors reveals their real-time trafficking. Neuron. 2005;48:977–985. - PubMed
-
- Bredt DS, Nicoll RA. AMPA receptor trafficking at excitatory synapses. Neuron. 2003;40:361–379. - PubMed
-
- Collingridge GL, Isaac JT, Wang YT. Receptor trafficking and synaptic plasticity. Nat Rev Neurosci. 2004;5:952–962. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
