

Endocannabinoids shape accumbal encoding of cue-motivated behavior via CB1 receptor activation in the ventral tegmentum
- PMID: 22284189
- PMCID: PMC3269037
- DOI: 10.1016/j.neuron.2011.11.018
Endocannabinoids shape accumbal encoding of cue-motivated behavior via CB1 receptor activation in the ventral tegmentum
Abstract
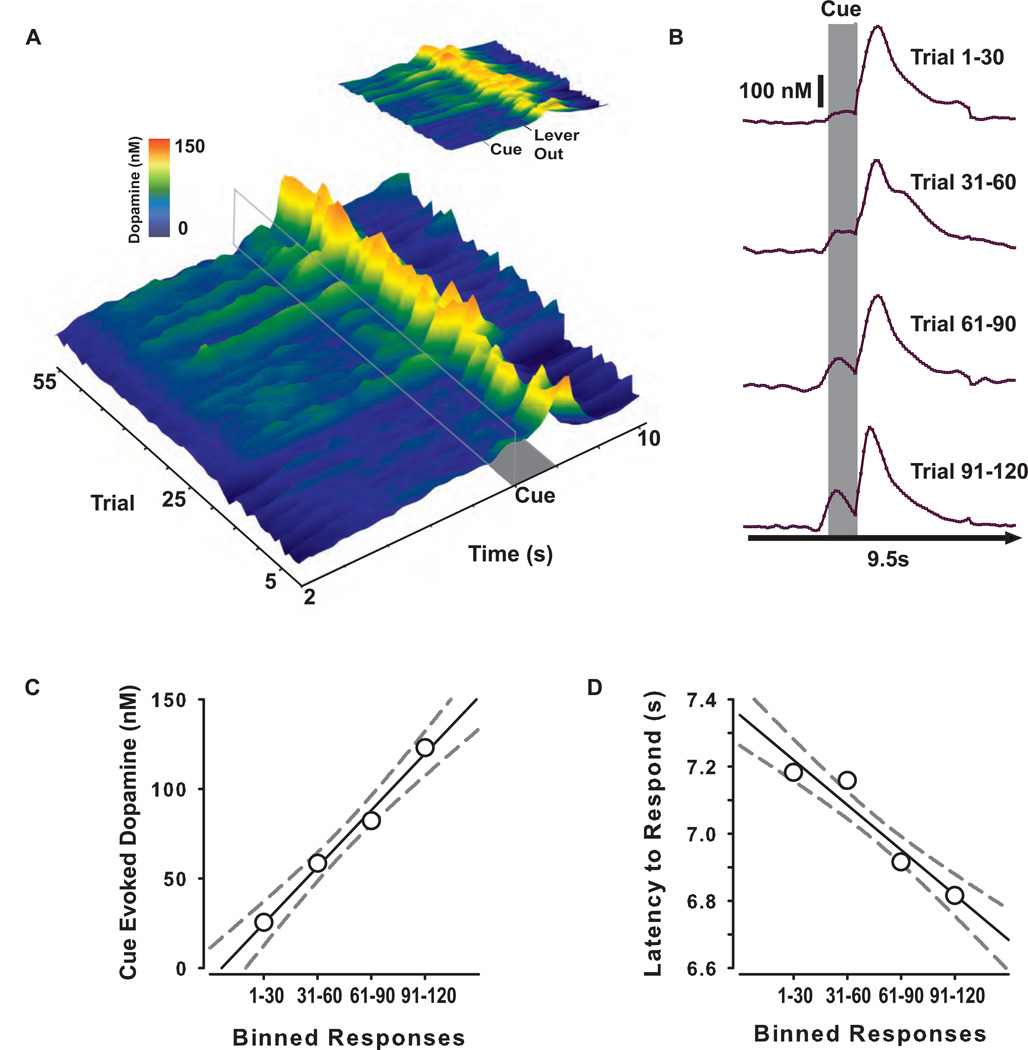
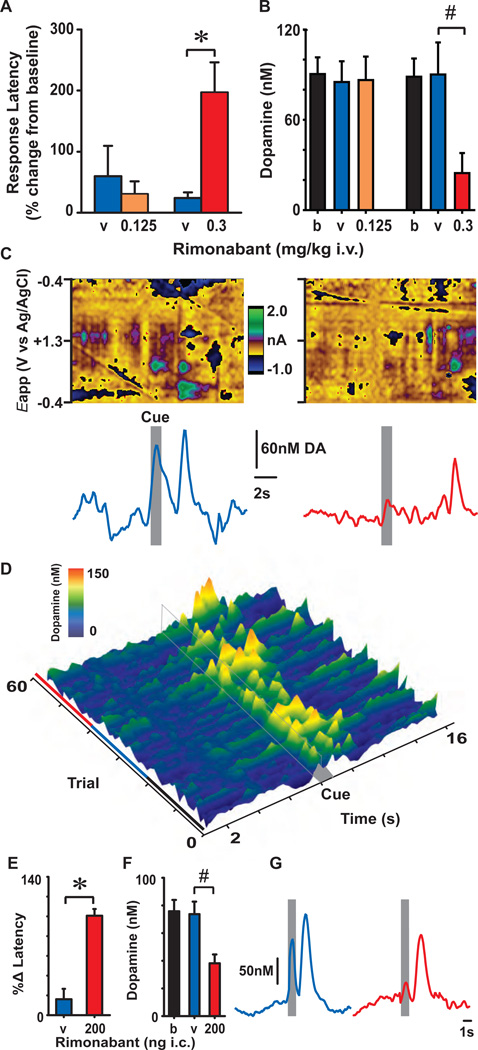
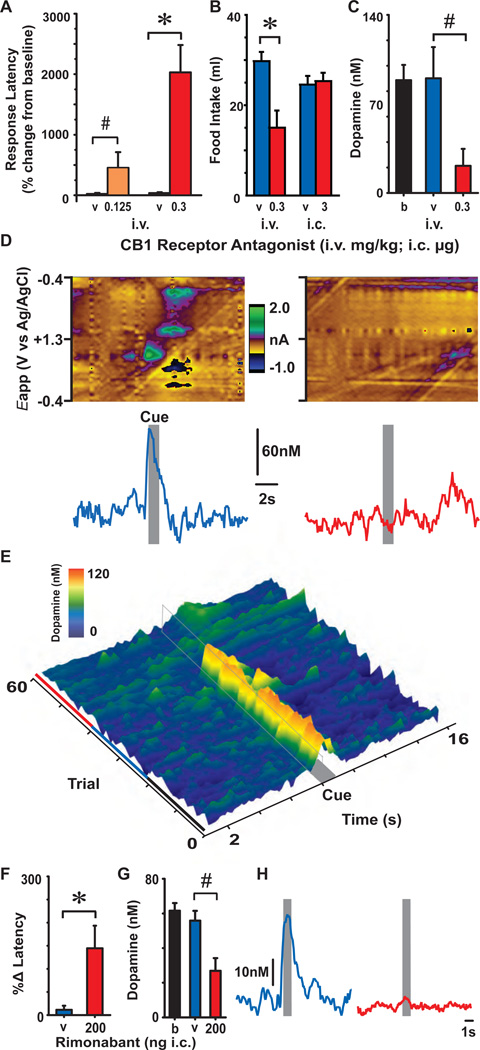
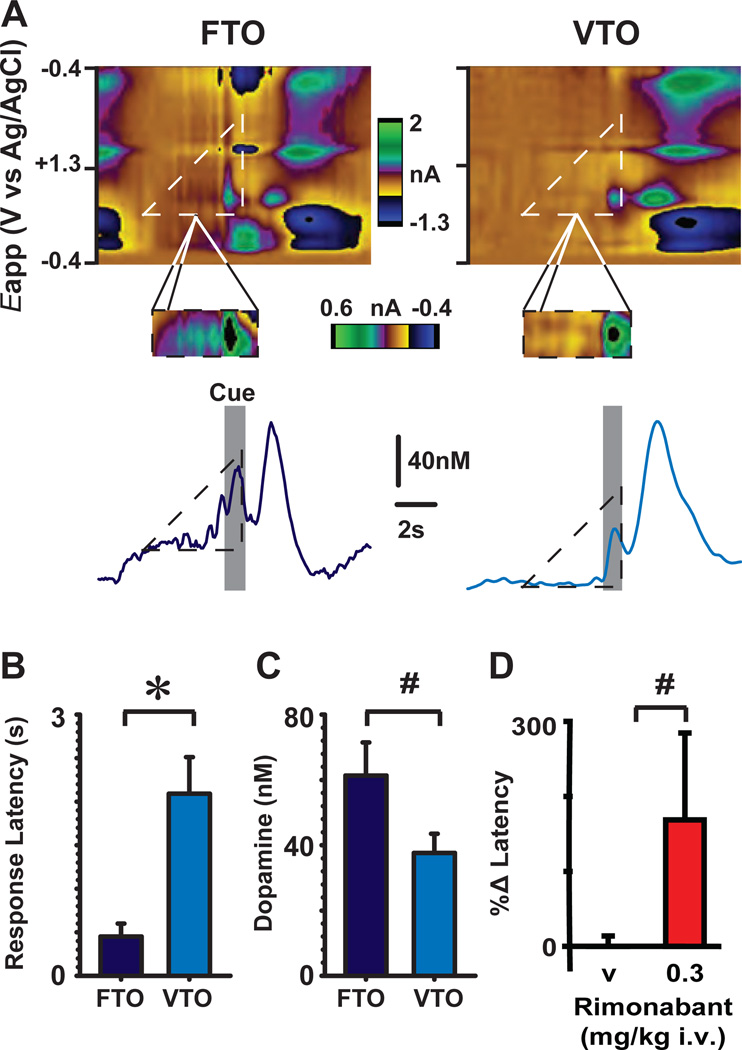
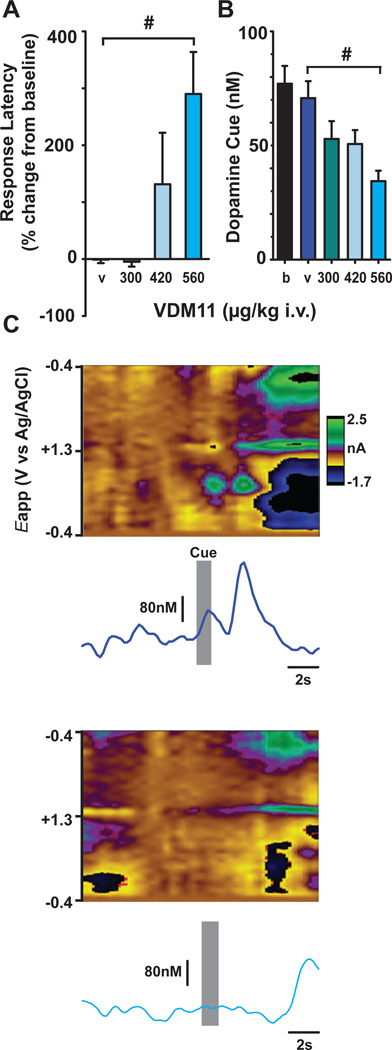
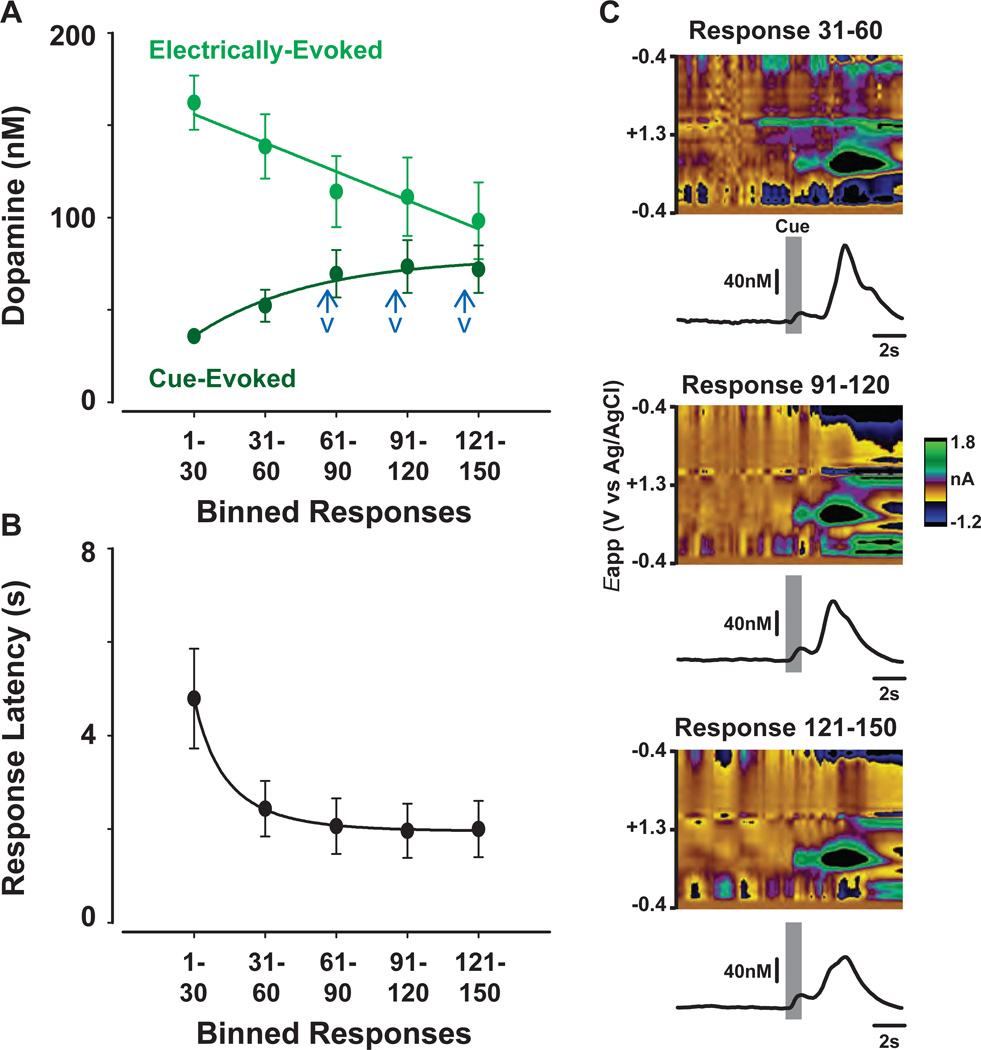
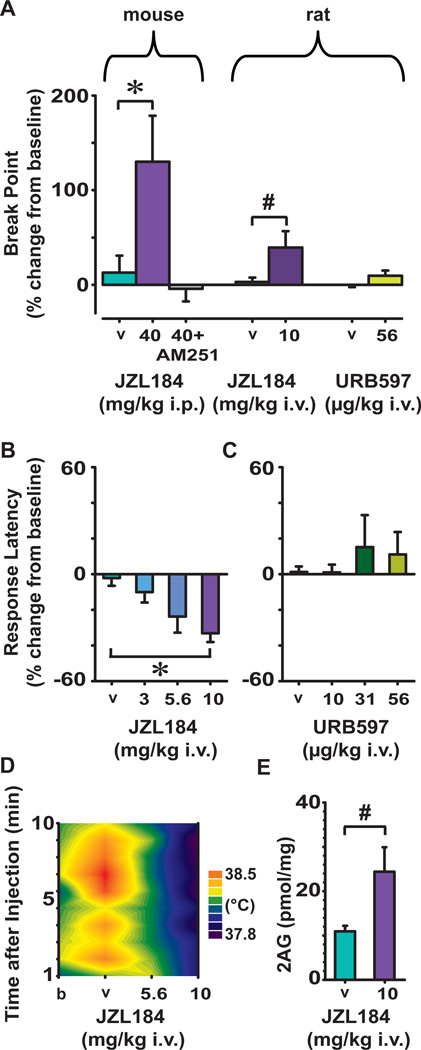
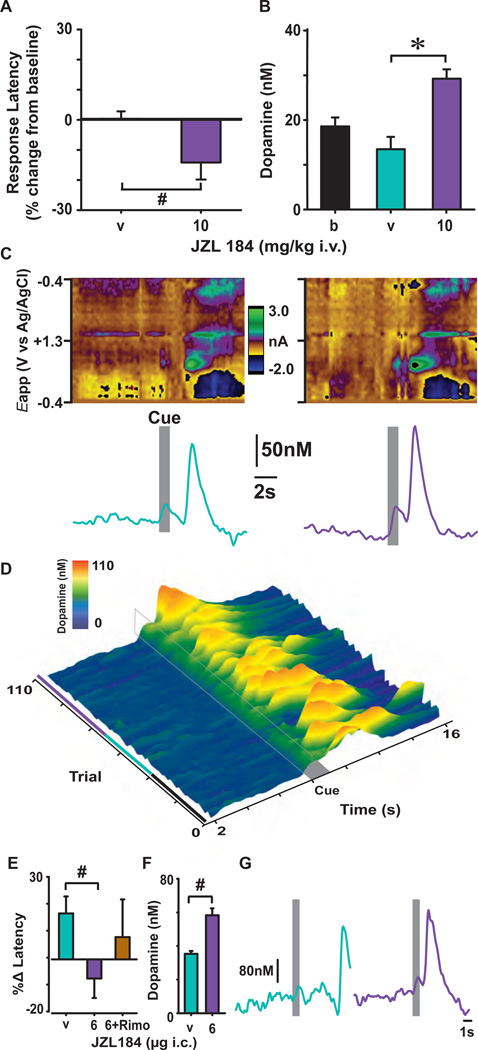
Transient increases in nucleus accumbens (NAc) dopamine concentration are observed when animals are presented with motivationally salient stimuli and are theorized to energize reward seeking. They arise from high-frequency firing of dopamine neurons in the ventral tegmental area (VTA), which also results in the release of endocannabinoids from dopamine cell bodies. In this context, endocannabinoids are thought to regulate reward seeking by modulating dopamine signaling, although a direct link has never been demonstrated. To test this, we pharmacologically manipulated endocannabinoid neurotransmission in the VTA while measuring transient changes in dopamine concentration in the NAc during reward seeking. Disrupting endocannabinoid signaling dramatically reduced, whereas augmenting levels of the endocannabinoid 2-arachidonoylglycerol (2AG) increased, cue-evoked dopamine concentrations and reward seeking. These data suggest that 2AG in the VTA regulates reward seeking by sculpting ethologically relevant patterns of dopamine release during reward-directed behavior.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Alvarez-Jaimes L, Polis I, Parsons LH. Attenuation of cue-induced heroin-seeking behavior by cannabinoid CB1 antagonist infusions into the nucleus accumbens core and prefrontal cortex, but not basolateral amygdala. Neuropsychopharmacology. 2008;33:2483–2493. - PubMed
-
- Berridge KC, Robinson TE. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Brain Res Rev. 1998;28:309–369. - PubMed
-
- Bindra D. Neuropsychological Interpretation of the Effects of Drive and Incentive-Motivation on General and Instrumental Behavior. Psychological Review. 1968;75:1–22.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
