

Review
doi: 10.1111/j.1742-4658.2012.08509.x.
Epub 2012 Feb 24.
Structural basis for protein phosphatase 1 regulation and specificity
Affiliations
- PMID: 22284538
- PMCID: PMC3350600
- DOI: 10.1111/j.1742-4658.2012.08509.x
Item in Clipboard
Review
Structural basis for protein phosphatase 1 regulation and specificity
FEBS J.
2013 Jan.
Abstract
The ubiquitous serine/threonine protein phosphatase 1 (PP1) regulates diverse, essential cellular processes such as cell cycle progression, protein synthesis, muscle contraction, carbohydrate metabolism, transcription and neuronal signaling. However, the free catalytic subunit of PP1, while an effective enzyme, lacks substrate specificity. Instead, it depends on a diverse set of regulatory proteins (≥ 200) to confer specificity towards distinct substrates. Here, we discuss recent advances in structural studies of PP1 holoenzyme complexes and summarize the new insights these studies have provided into the molecular basis of PP1 regulation and specificity.
© 2012 The Authors Journal compilation © 2012 FEBS.
Figures
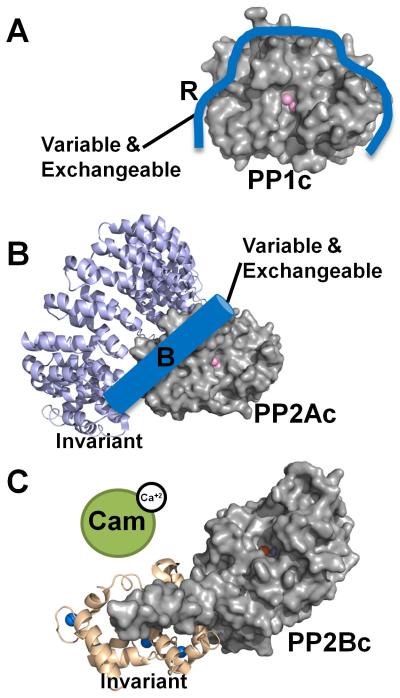
PP1, PP2A and PP2B (Calcineurin, PP3) are metallo-enzymes with a highly similar and conserved catalytic domain structure. However, regulation is achieved via different modes. (A) PP1 is a single catalytic domain (PP1c, gray surface, active site metal ions are depicted as pink spheres; from PDB 3EGG), which can bind to ≥200 biochemically confirmed regulators (blue; indicated by ‘R’). Using one or more of these regulators PP1 is converted into hundreds of highly specific holoenzymes. (B) PP2A (PP2Ac, gray surface, active site metal ions are depicted as pink spheres; from PDB 2NYM) is regulated in a similar manner as PP1, as it binds to a variety of variable and exchangeable B-subunits (blue cylinder). It always binds to an invariant A subunit (magenta ribbon, from PDB 2NYM). The total number of achievable PP2A holoenzymes is ~75. (C) PP2B also consists of a catalytic A and an invariant structural B subunit (PP2Bc, gray surface, active site metal ions are depicted as orange and dark red spheres; B-subunit wheat ribbons, calcium ions are depicted as blue spheres; from PDB 2P6B). However, in order to become activated (release of the auto-inhibitory helix from the active site), calcium activated Calmodulin (green) must bind to PP2B.
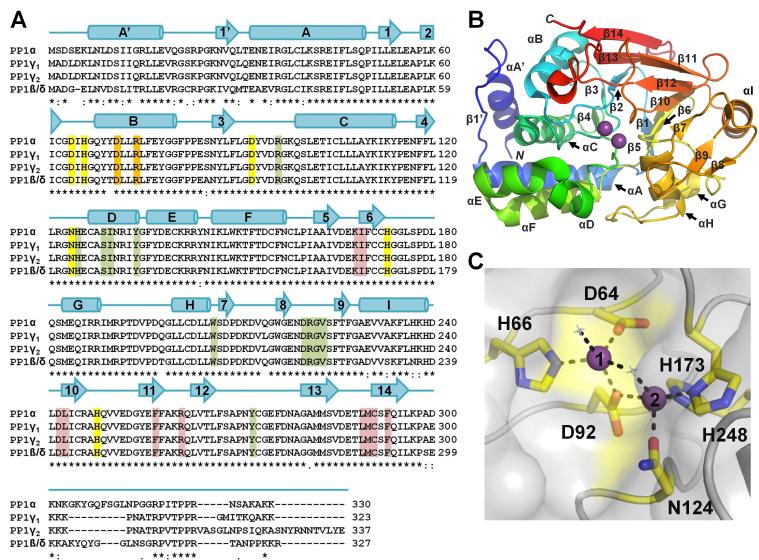
(A) Sequence alignment of PP1 α, β, γ1 and γ2 [all human]. Identical (*), conservative (:) and similar (.) residues are indicated. Sequence identity between corresponding isoforms is 93% (γ1/γ2), 91% (α/γ1), 89% (α/β), 88% (α/γ2), 87% (γ1/β), and 85% (γ2/β). Secondary structure elements of PP1α are indicated above the sequences with α-helices represented as cylinders and β-strands indicated with arrows. Metal coordination residues are highlighted in yellow, residues critical for coordinating molecular toxins are highlight in green (highlighted residues are within 5 Å of the bound toxin in each of the eight PP1:molecular toxin structures determined to date, see Table 1), residues that form the RVxF binding pocket are highlight in red and residues that play a central role in binding targeting proteins/substrates that bind in the PP1 C-terminal groove are highlighted in orange. (B) PP1α (3E7A) illustrated as a cartoon and colored from blue to red from the N- to the C-terminus, respectively. Bound metal ions are shown as spheres (magenta). PP1 secondary structural elements are labeled. (C) PP1α (3E7A) in a surface representation with the residues that coordinate the metals shown as sticks and colored yellow. Metals 1 and 2 are represented as purple spheres and numbered. Metal binding residues are 100% conserved in PP1, PP2A and PP2B.
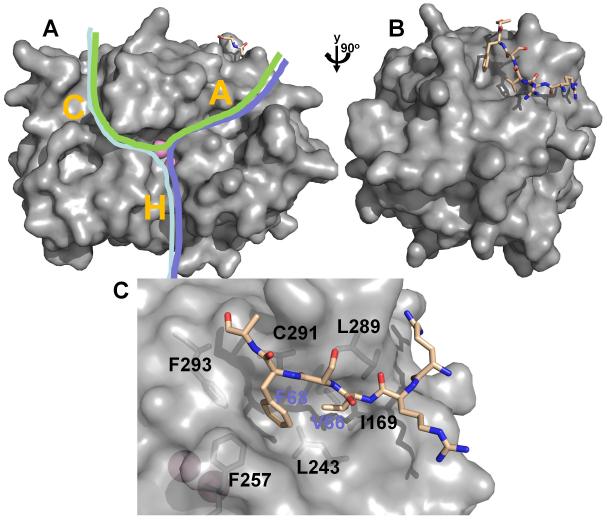
(A) PP1 (from the Gm:PP1 structure [38]) is shown as surface representation. Active site metal ions are depicted as pink spheres and the Gm RVxF peptide is shown in stick representation (beige). The C-terminal (C), hydrophobic (H) and acidic (A) substrate binding grooves are indicated with a letter. Different PP1 substrates (indicated by green, pink and blue lines) can bind via a single groove, or, as shown here, multiple grooves, illustrating that substrates likely bind PP1 via a variety of mechanisms. (B) PP1 is rotated by 90° in order to highlight the RVxF peptide binding site. (C) Detailed view of the RVxF binding site, illustrating the deep hydrophobic pockets which bind the ‘V’ and ‘F’ residues (V66 and F68 for the Gm peptide) of the RVxF motif.
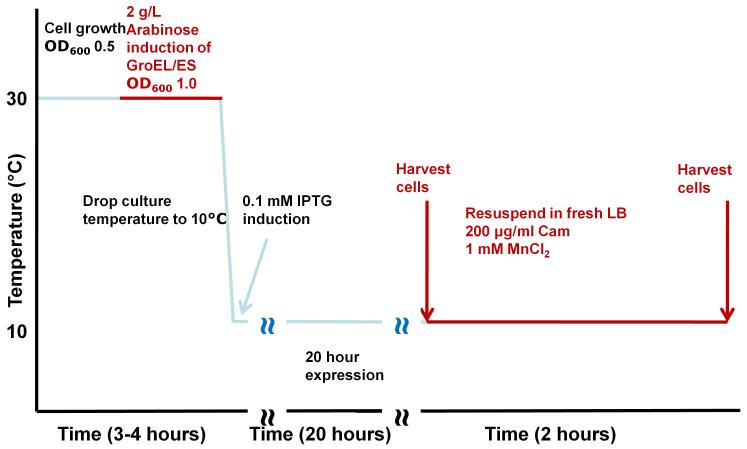
Detailed scheme (timeline) illustrating the steps of our recently developed PP1 expression protocol.
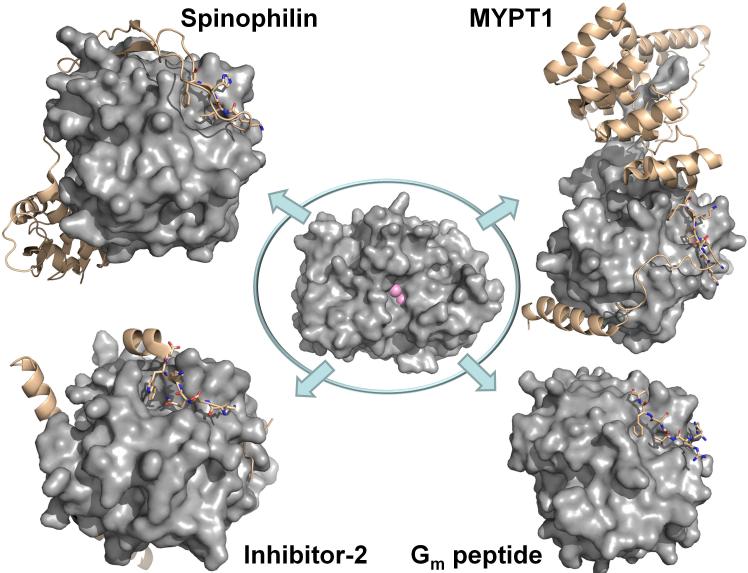
Center: PP1 is a single catalytic domain (PP1c, gray surface, active site metal ions are depicted as pink spheres; from PDB 3EGG). Upper left. The spinophilin:PP1 holoenzyme (spinophilin is shown as beige ribbons; RVxF residues are highlighted as sticks. Lower left. The I-2:PP1 holoenzyme (I-2 is shown as beige ribbons; RVxF residues are highlighted as sticks). Upper right. The MYPT1:PP1 holoenzyme (MYPT1 is shown as beige ribbons; RVxF residues are highlighted as sticks). Lower right. The Gm:PP1 RVxF peptide structure (the RVxF peptide is shown as beige sticks).
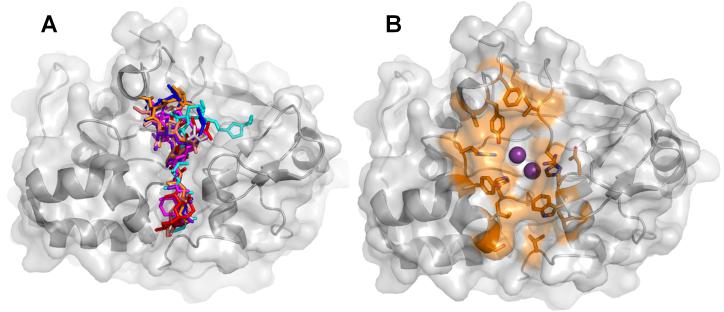
(A) Overlay of the eight PP1:toxin complexes; toxins are illustrated as sticks (3E7A, grey; 3E7B, red; 1IT6, cyan; 1JK7, magenta; 2BCD, salmon; 3EGH, purple, 1FMJ, blue; 2BDX, orange). (B) The PP1 residues that mediate toxin binding. Highlighted residues (shown as sticks and colored orange) are within 5 Å of the bound toxin in each of the PP1:molecular toxin structures determined to date.
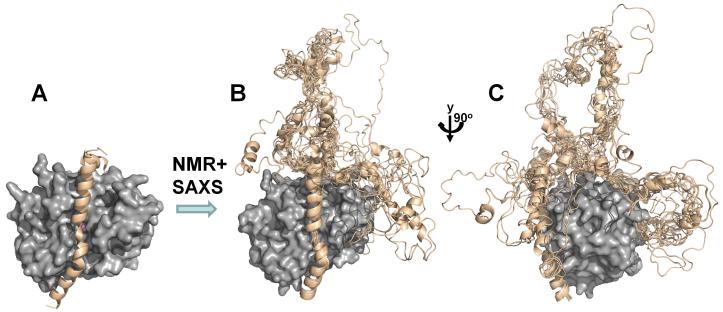
(A) I-2:PP1 holoenzyme X-ray structure (PP1c, gray surface, I-2 beige ribbons, PDB 2O8G); (B) the use of NMR and SAXS allowed for the determination of the I-2:PP1 ensemble structure; (C) PP1 is rotated by 90° in order to highlight the I-2 RVxF and SILK peptide binding sites.
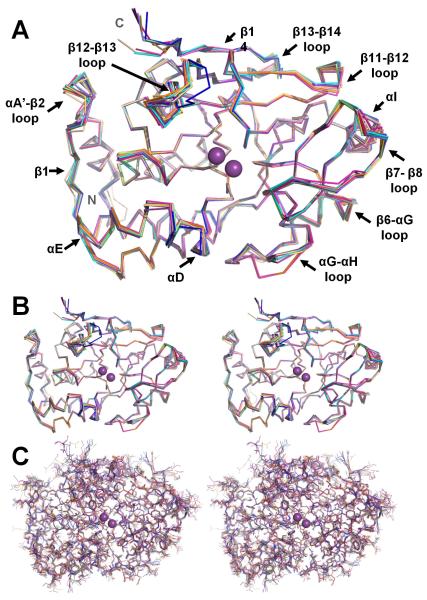
(A) Overlay of the PP1 structures, illustrated as backbone ribbons (1FMJ, dark blue; 1IT6, cyan; 1JK7, light magenta; 1S70, beige; 2BCD, salmon; 2BDX, orange; 2O8G, slate; 3E7A, grey; 3E7B, lime green; 2EGG teal; 3HVQ, pink; 3N5U, yellow-orange; 3EGH, purple-blue; Gm, violet-purple). The bound metals are represented as spheres and colored magenta. The regions of PP1 that undergo the changes in conformation between the structures are labeled. (B) Stereo view of the overlay in A. (C) Same stereo view as in B except all atoms are shown.
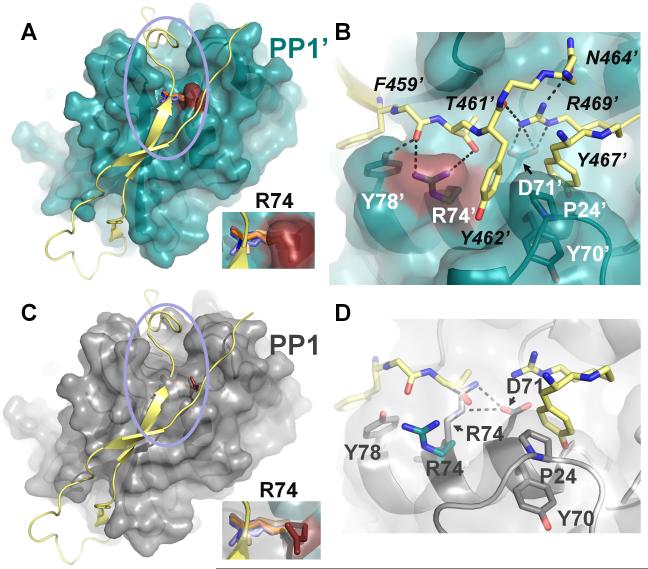
(A) PP1:spinophilin complex (3EGG). Spinophilin (yellow) is shown in cartoon representation. PP1 (teal) is illustrated as a transparent surface, with R74 colored in red. Residue R74 from all PP1 structures determined to date are shown in stick representation (with the exception of the PP1:spinophilin:nodularin and PP1:neurabin complexes, which are identical to the PP1:spinophilin complex and not included for clarity). PP1 structures were superimposed as in Figure 1. In all cases, with the exception of the PP1:spinophilin complex, R74 lies across the C-terminal groove. Inset, close-up of R74. (B) Close-up of the area enclosed in the pink circle in (A). Colors identical to those in (A) with residues numbers for PP1 (white) and spinophilin (black) indicated (spinophilin residues 465 and 466 were removed for clarity, as were some carbonyls). Polar interactions between PP1 side chains and spinophilin are indicated by black dashed lines. Spinophilin residue R469 makes multiple electrostatic contacts with PP1 residue Asp71 and other spinophilin residues. (C) Same as (A) except PP1 (grey) is from the PP1:nodularin complex (3E7A). (D) Same as (B) except PP1 residues (grey) are now from PP1:nodularin complex. The polar interactions between PP1 residues R74 and D71 are illustrated by dashed lines (black). R74 (teal) from the PP1:spinophilin complex is also shown to illustrate the large conformational change of this residue between the two complexes. Finally, a subset of the spinophilin residues in (B) show that the side chain of spinophilin T461 sterically clashes that of PP1:nodularin complex PP1 residue R74, explaining why R74 must change conformation in order for spinophilin to bind PP1.
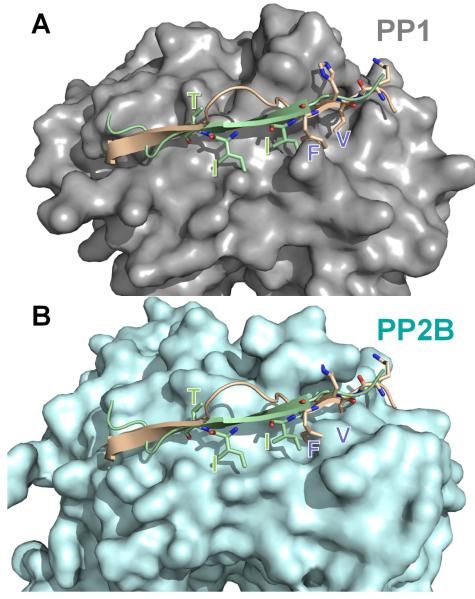
The spinophilin:PP1 (PDB 3EGG) holoenzyme was superimposed onto PP2B:PIVIIT (PDB 2P6B) using PP1 and PP2B. In (A), PP1 (grey) is shown in surface representation with spinophilin (residues 446-461 of spinophilin, which includes the RVxF motif and the β14 binding residues) in beige and the PIVIIT peptide in green. As can be seen, both the PP1 regulator spinophilin, as well as the PP2B substrate mimicking PIVIIT peptide bind to the β14 sheet of PP1/PP2B. (B) Same as in ‘A’ except PP2B (light blue) is now shown in surface representation. The residues that are essential for mediating phosphatase specific interactions are shown in stick representation and labeled in both panels. Comparison of panels ‘A’ and ‘B’ illustrates that the deep hydrophobic pockets that are present in PP1 (RVxF binding pockets) or PP2B (PIVIIT binding pockets) are not present in the other phosphatase. This structural data provides the first molecular insight into the mechanism of specificity regulation between the highly homologous enzymes PP1 and PP2B.
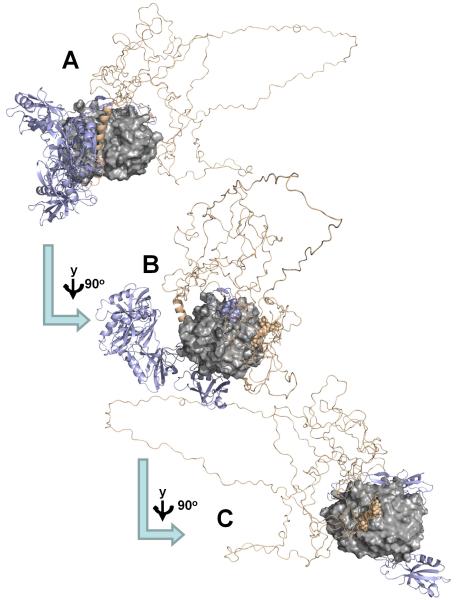
(A) SAXS-based ensemble structure for the PP1:spinophilin:I-2 (PSI) complex, which highlights the increased flexibility of I-2 in the PSI complex versus the I-2:PP1 complex (compare with Fig. 5). (PP1c, gray surface; I-2, wheat ribbon, spinophilin, slate ribbons); (B) PP1 is rotated by 90° in order to highlight the spinophilin RVxF peptide binding site (spinophilin residues I449 and F451, as part of the RVxF motif, are highlighted as spheres); (C) PP1 is rotated by additional 90° in order to highlight the I-2 SILK peptide binding site (I-2 residues 13-16, the SILK motif, are highlighted as spheres).
References
-
- Olsen JV, Vermeulen M, Santamaria A, Kumar C, Miller ML, Jensen LJ, Gnad F, Cox J, Jensen TS, Nigg EA, Brunak S, Mann M. Quantitative phosphoproteomics reveals widespread full phosphorylation site occupancy during mitosis. Sci Signal. 2010;3:ra3. - PubMed
-
- Shi Y. Serine/threonine phosphatases: mechanism through structure. Cell. 2009;139:468–84. - PubMed
-
- Patterson KI, Brummer T, O’Brien PM, Daly RJ. Dual-specificity phosphatases: critical regulators with diverse cellular targets. Biochem J. 2009;418:475–89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
