

Selective binding of anti-DNA antibodies to native dsDNA fragments of differing sequence
- PMID: 22285306
- PMCID: PMC3655556
- DOI: 10.1016/j.imlet.2012.01.003
Selective binding of anti-DNA antibodies to native dsDNA fragments of differing sequence
Abstract
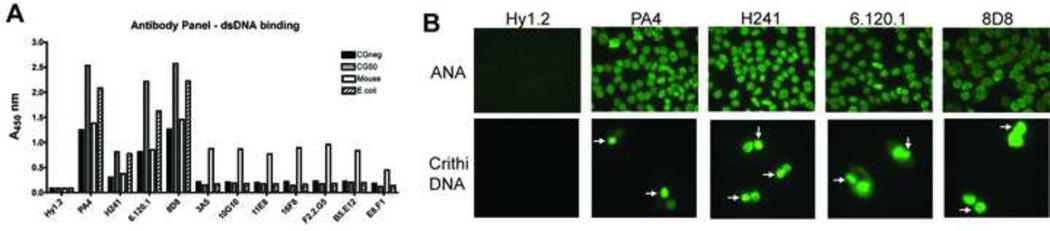
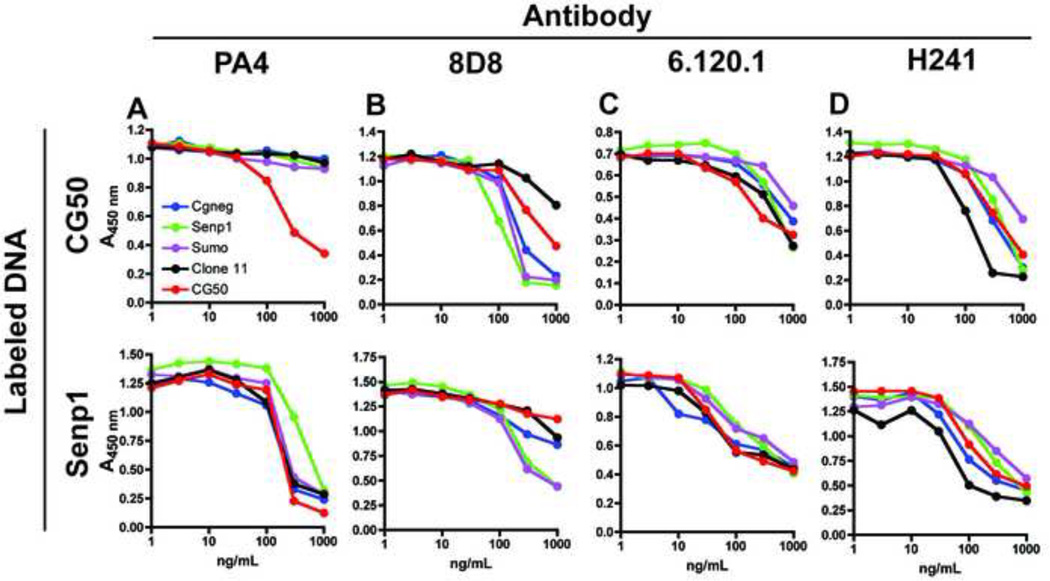
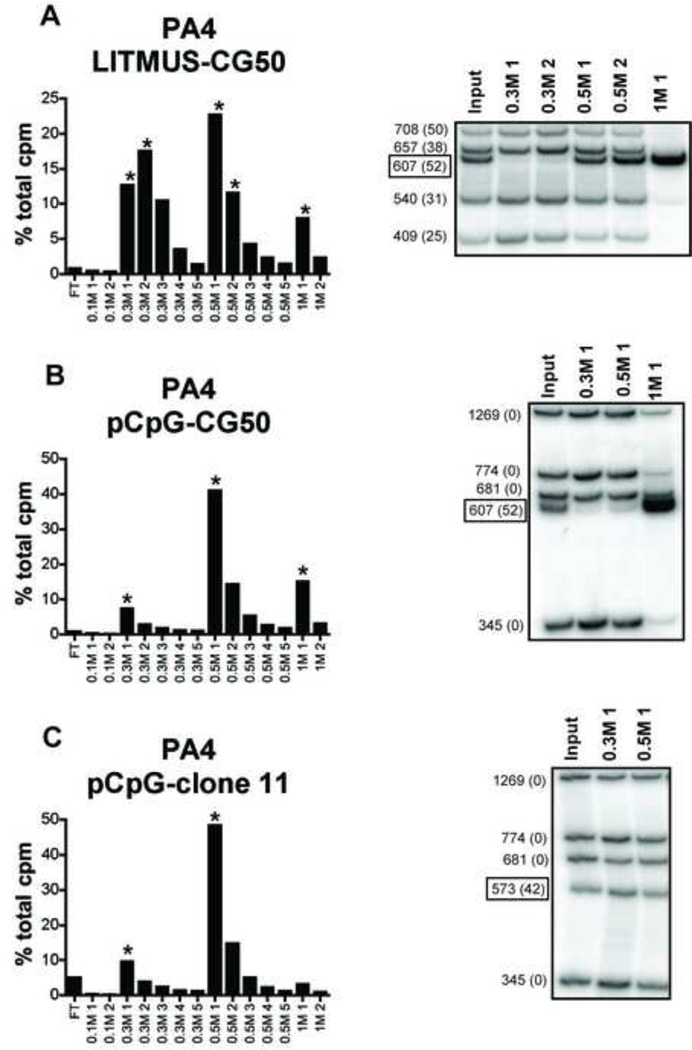
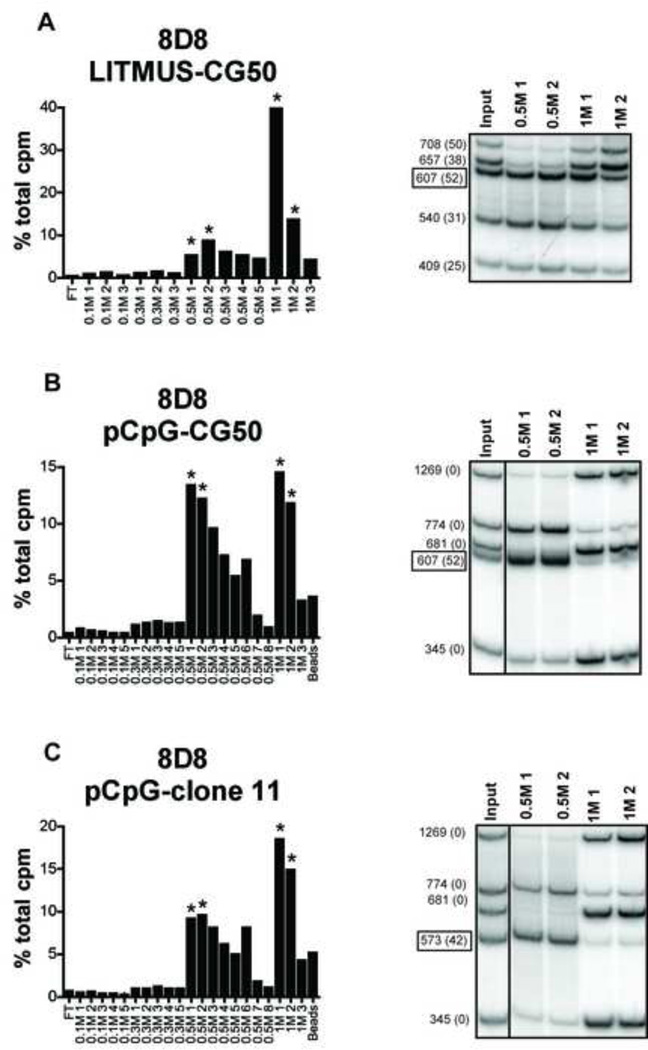
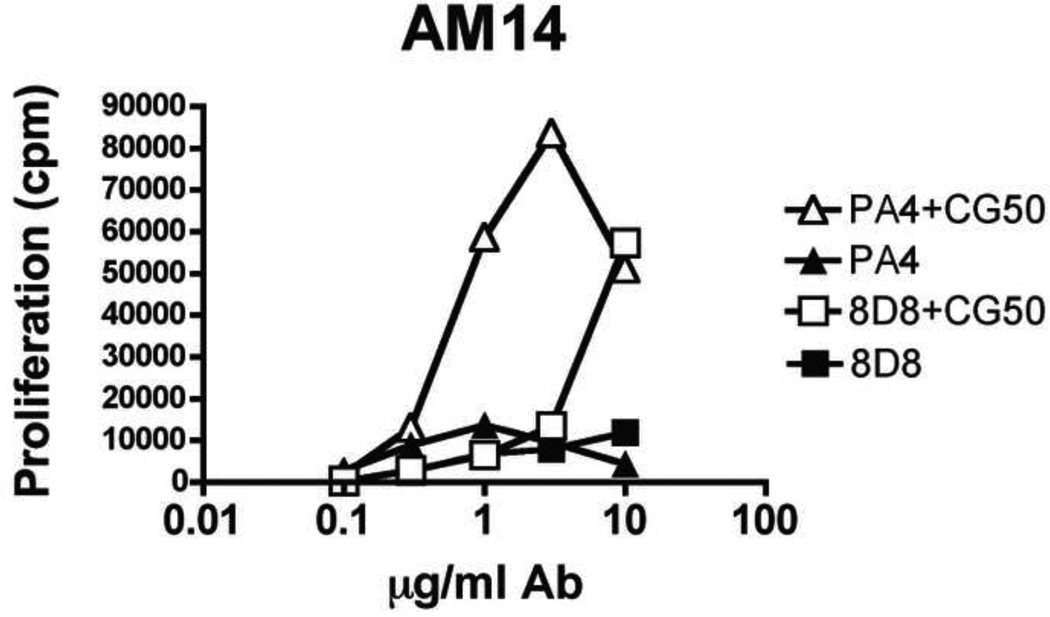
Systemic autoimmune diseases are characterized by the development of autoantibodies directed against a limited subset of nuclear antigens, including DNA. DNA-specific B cells take up mammalian DNA through their B cell receptor, and this DNA is subsequently transported to an endosomal compartment where it can potentially engage TLR9. We have previously shown that ssDNA-specific B cells preferentially bind to particular DNA sequences, and antibody specificity for short synthetic oligodeoxynucleotides (ODNs). Since CpG-rich DNA, the ligand for TLR9 is found in low abundance in mammalian DNA, we sought to determine whether antibodies derived from DNA-reactive B cells showed binding preference for CpG-rich native dsDNA, and thereby select immunostimulatory DNA for delivery to TLR9. We examined a panel of anti-DNA antibodies for binding to CpG-rich and CpG-poor DNA fragments. We show that a number of anti-DNA antibodies do show preference for binding to certain native dsDNA fragments of differing sequence, but this does not correlate directly with the presence of CpG dinucleotides. An antibody with preference for binding to a fragment containing optimal CpG motifs was able to promote B cell proliferation to this fragment at 10-fold lower antibody concentrations than an antibody that did not selectively bind to this fragment, indicating that antibody binding preference can influence autoreactive B cell responses.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Plotz PH. The autoantibody repertoire: searching for order. Nat Rev Immunol. 2003;3:73–78. - PubMed
-
- Leadbetter EA, Rifkin IR, Hohlbaum AM, Beaudette BC, Shlomchik MJ, Marshak- Rothstein A. Chromatin-IgG complexes activate B cells by dual engagement of IgM and Toll-like receptors. Nature. 2002;416:603–607. - PubMed
-
- Shlomchik MJ, Zharhary D, Saunders T, Camper SA, Weigert MG. A rheumatoid factor transgenic mouse model of autoantibody regulation. Int Immunol. 1993;5:1329–1341. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
