

Chromatin dynamics in DNA double-strand break repair
- PMID: 22285574
- PMCID: PMC3368990
- DOI: 10.1016/j.bbagrm.2012.01.002
Chromatin dynamics in DNA double-strand break repair
Abstract
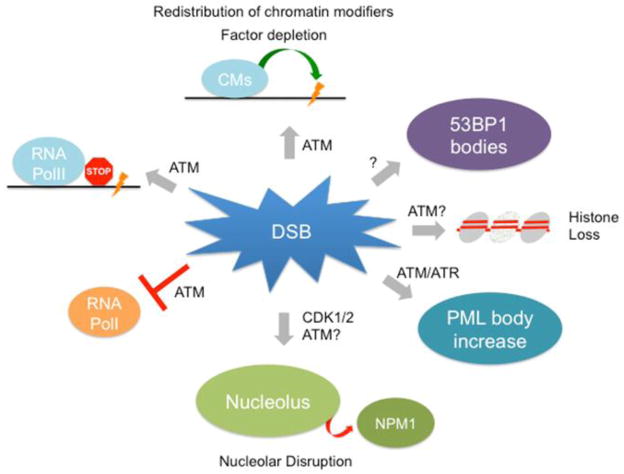
DNA double-strand breaks (DSBs) occur in the context of a highly organized chromatin environment and are, thus, a significant threat to the epigenomic integrity of eukaryotic cells. Changes in break-proximal chromatin structure are thought to be a prerequisite for efficient DNA repair and may help protect the structural integrity of the nucleus. Unlike most bona fide DNA repair factors, chromatin influences the repair process at several levels: the existing chromatin context at the site of damage directly affects the access and kinetics of the repair machinery; DSB induced chromatin modifications influence the choice of repair factors, thereby modulating repair outcome; lastly, DNA damage can have a significant impact on chromatin beyond the site of damage. We will discuss recent findings that highlight both the complexity and importance of dynamic and tightly orchestrated chromatin reorganization to ensure efficient DSB repair and nuclear integrity. This article is part of a Special Issue entitled: Chromatin in time and space.
Published by Elsevier B.V.
Figures

References
-
- Talbert PB, Henikoff S. Histone variants--ancient wrap artists of the epigenome. Nat Rev Mol Cell Biol. 2010;11:264–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
