

Computational modeling of cellular signaling processes embedded into dynamic spatial contexts
- PMID: 22286385
- PMCID: PMC3448286
- DOI: 10.1038/nmeth.1861
Computational modeling of cellular signaling processes embedded into dynamic spatial contexts
Abstract
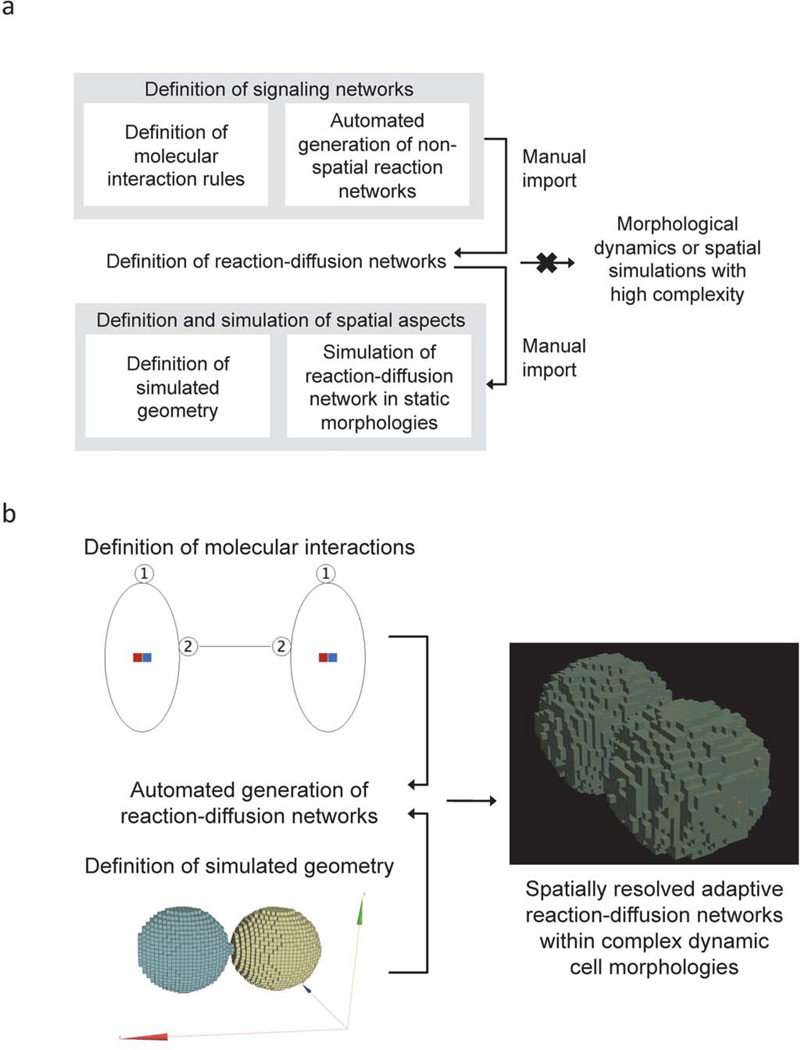
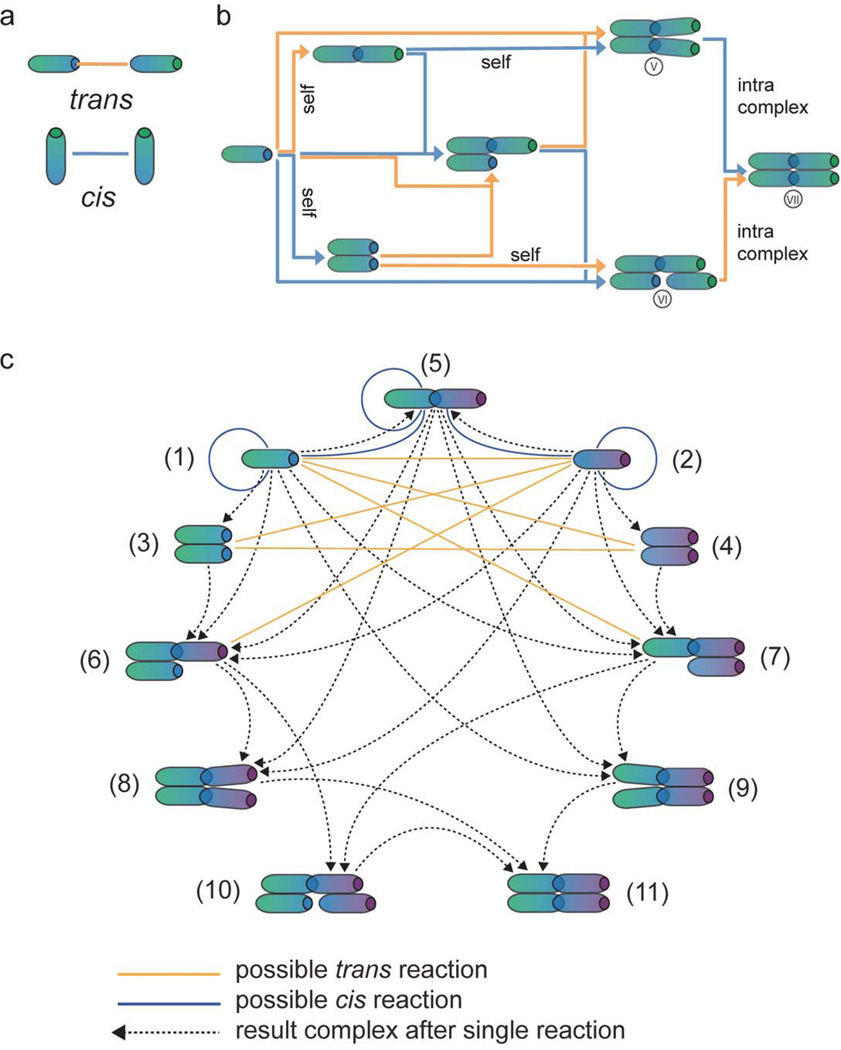
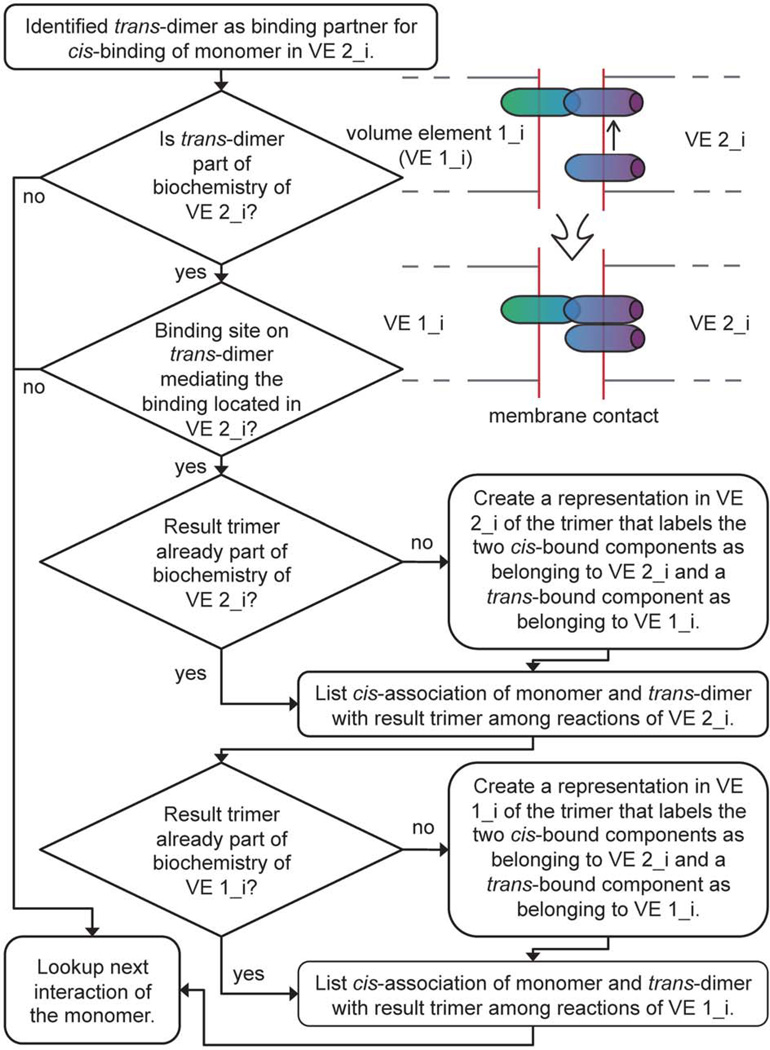
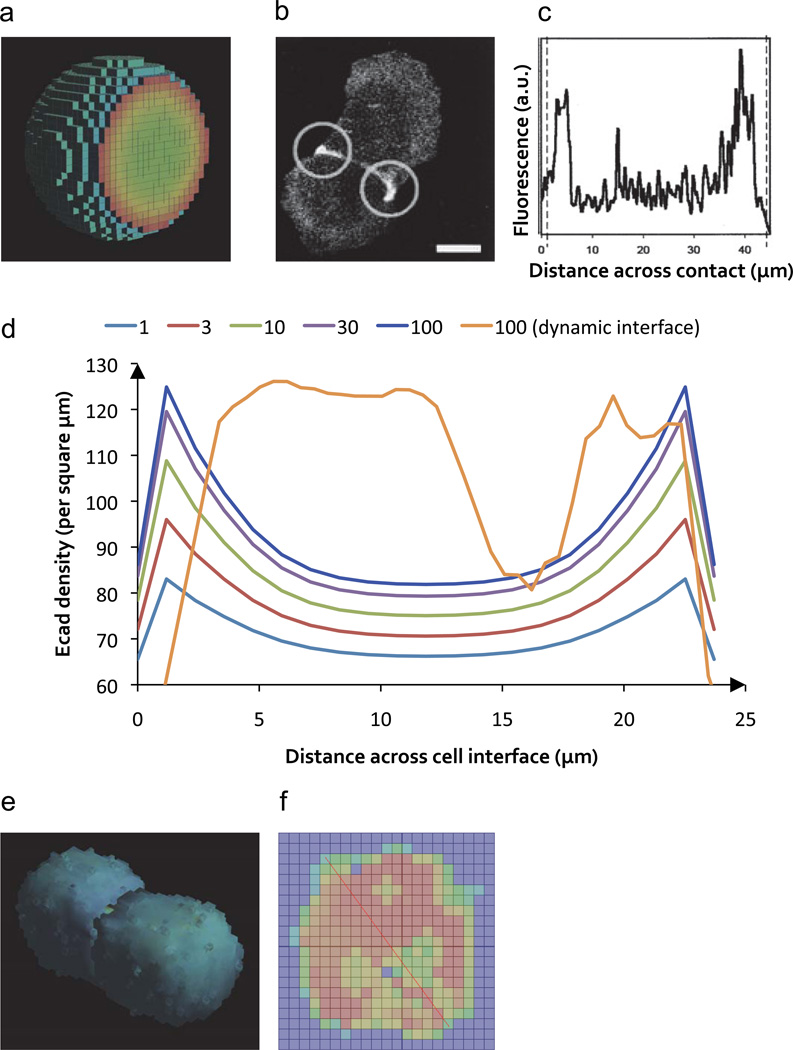
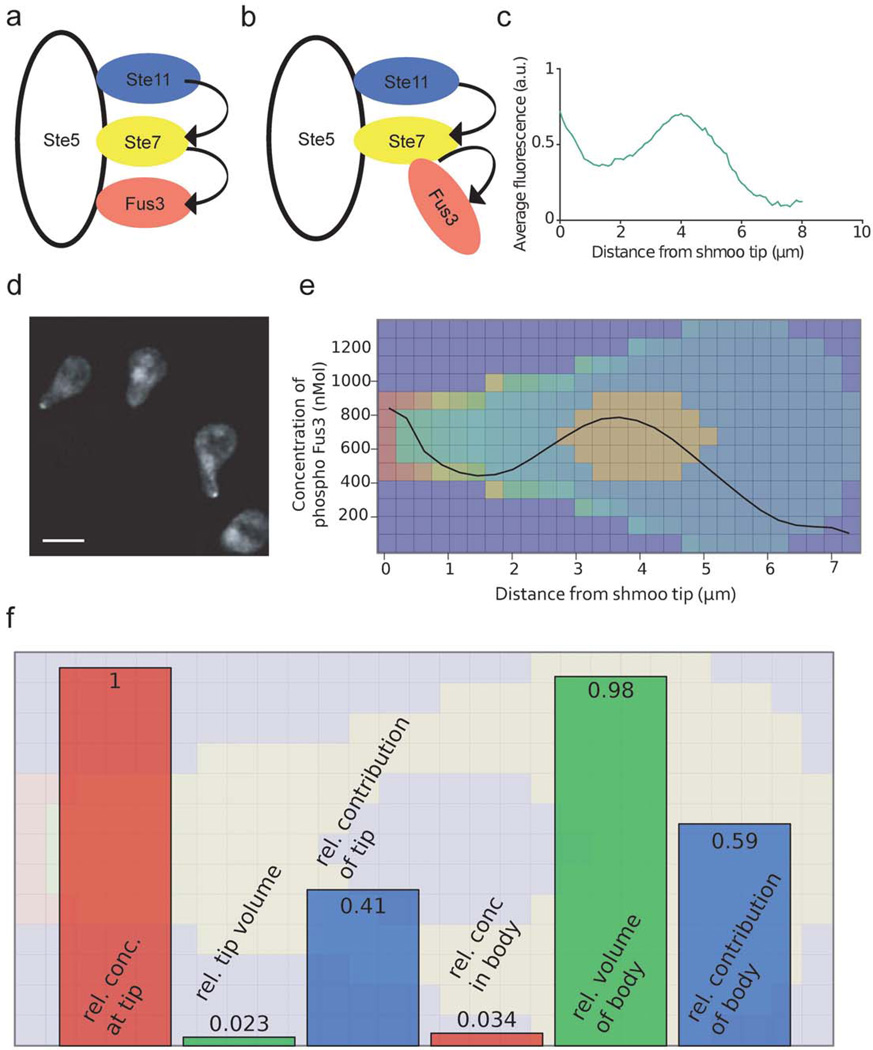
Cellular signaling processes depend on spatiotemporal distributions of molecular components. Multicolor, high-resolution microscopy permits detailed assessment of such distributions, providing input for fine-grained computational models that explore mechanisms governing dynamic assembly of multimolecular complexes and their role in shaping cellular behavior. However, it is challenging to incorporate into such models both complex molecular reaction cascades and the spatial localization of signaling components in dynamic cellular morphologies. Here we introduce an approach to address these challenges by automatically generating computational representations of complex reaction networks based on simple bimolecular interaction rules embedded into detailed, adaptive models of cellular morphology. Using examples of receptor-mediated cellular adhesion and signal-induced localized mitogen-activated protein kinase (MAPK) activation in yeast, we illustrate the capacity of this simulation technique to provide insights into cell biological processes. The modeling algorithms, implemented in a new version of the Simmune toolset, are accessible through intuitive graphical interfaces and programming libraries.
Figures
References
-
- Lingwood D, Simons K. Lipid rafts as a membrane-organizing principle. Science. 2010;327:46–50. - PubMed
-
- Kholodenko BN. Four-dimensional organization of protein kinase signaling cascades: the roles of diffusion, endocytosis and molecular motors. J Exp Biol. 2003;206:2073–2082. - PubMed
-
- Delon J, Germain RN. Information transfer at the immunological synapse. Curr Biol. 2000;10:R923–R933. - PubMed
-
- Jones RB, Gordus A, Krall JA, MacBeath G. A quantitative protein interaction network for the ErbB receptors using protein microarrays. Nature. 2006;439:168–174. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
