

The PKCθ pathway participates in the aberrant accumulation of Fra-1 protein in invasive ER-negative breast cancer cells
- PMID: 22286759
- PMCID: PMC3624663
- DOI: 10.1038/onc.2011.659
The PKCθ pathway participates in the aberrant accumulation of Fra-1 protein in invasive ER-negative breast cancer cells
Abstract
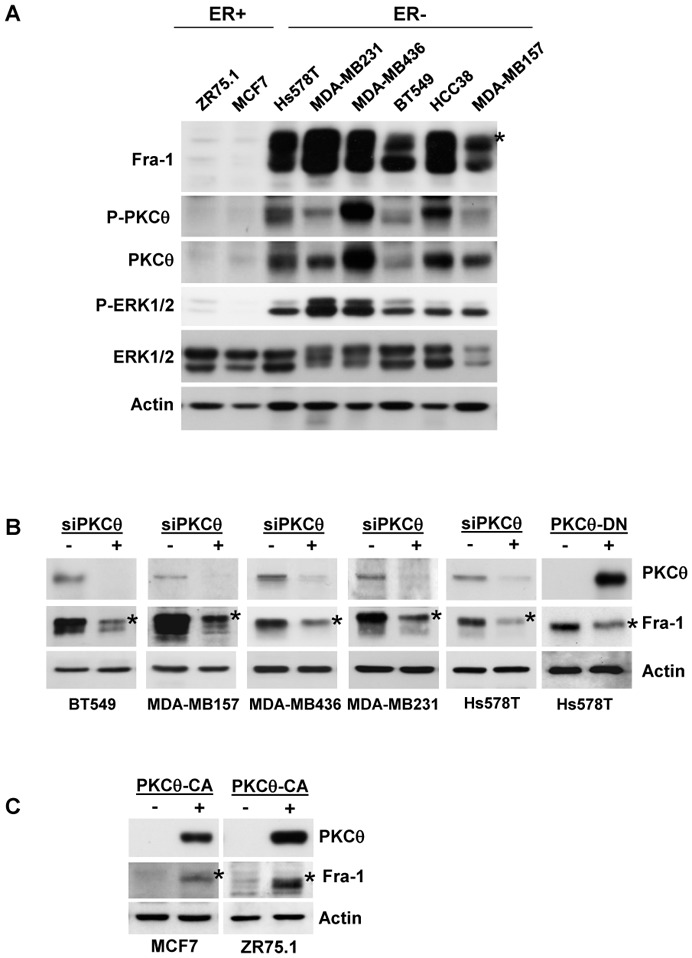
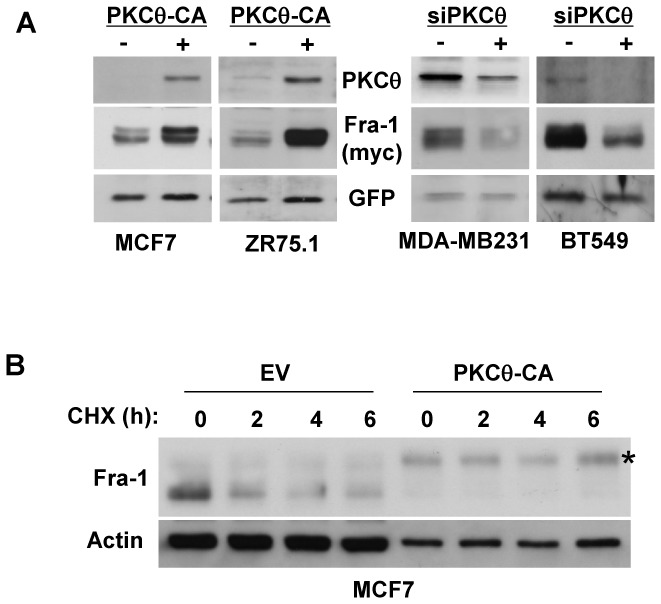
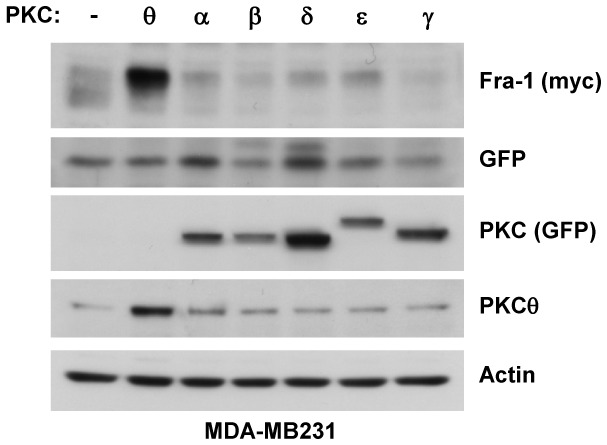
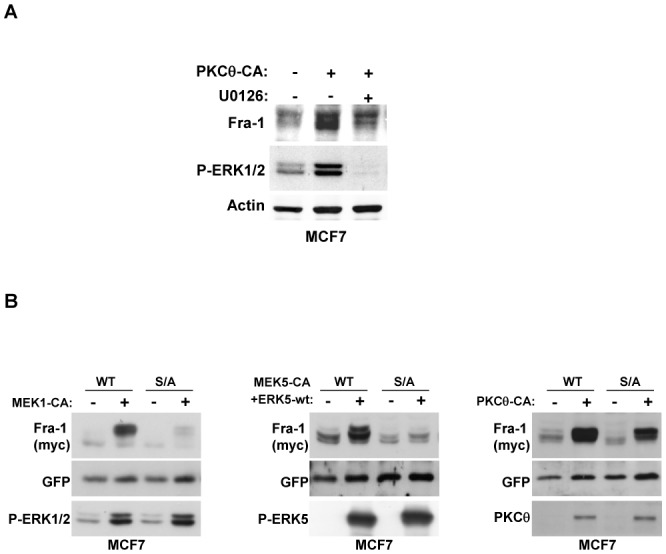
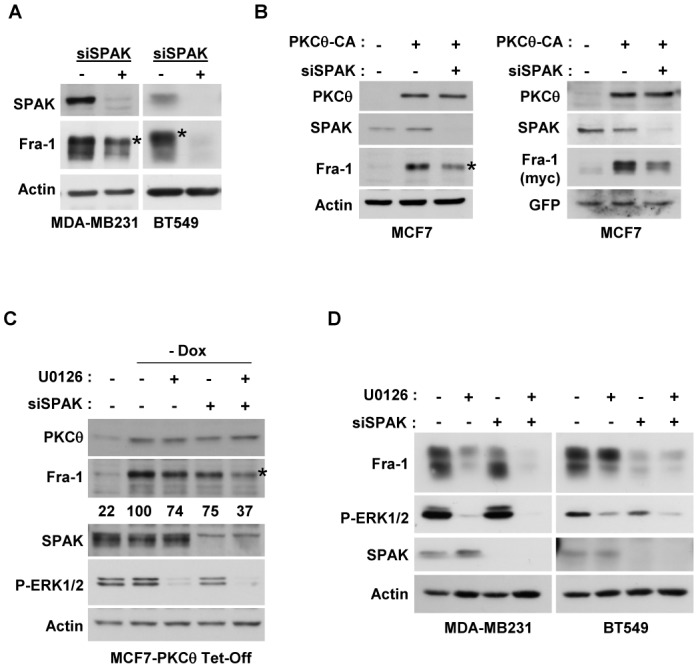
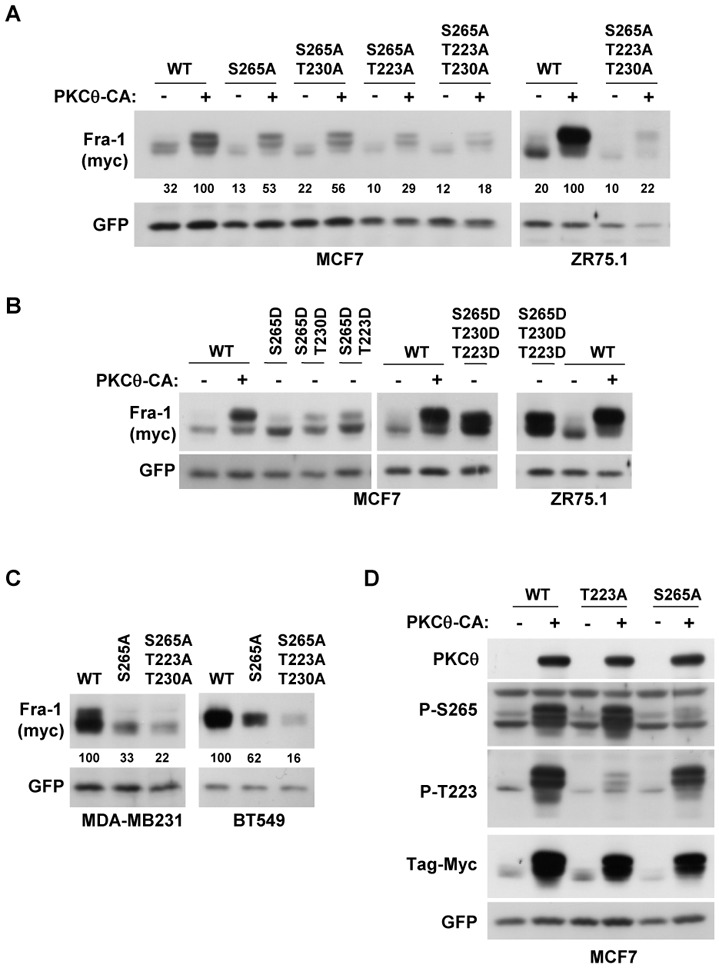
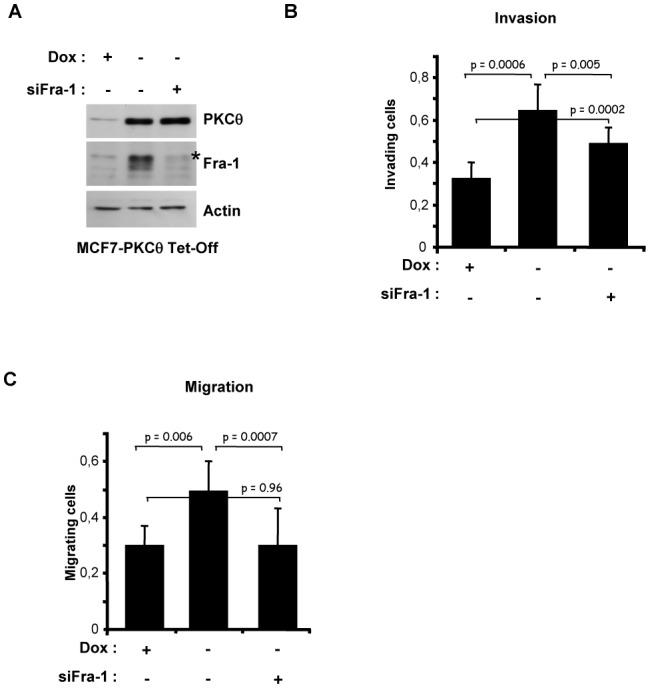
Fra-1 is aberrantly expressed in a large number of cancer cells and tissues, and emerging evidence suggests an important role for this Fos family protein in both oncogenesis and the progression or maintenance of many tumour types. Here, we show that the concentration of Fra-1 is high in invasive oestrogen receptor (ER)-negative (ER-) breast cancer cell lines, regardless of their Ras pathway status. All of the ER- cells express high levels of activated PKCθ, and the inhibition of PKCθ activity using RNA interference or the expression of a dominant-negative mutant results in a dramatic reduction in Fra-1 abundance. Conversely, the ectopic expression of constitutively active PKCθ leads to Fra-1 phosphorylation and accumulation in poorly invasive ER+ cells. This accumulation is due to the stabilisation of the Fra-1 protein through PKCθ signalling, whereas other members of the PKC family are ineffective. Both Ste20-related proline-alanine-rich kinase (SPAK) and ERK1/2, whose activities are upregulated by PKCθ, participate in PKCθ-driven Fra-1 stabilisation. Interestingly, their relative contributions appear to be different depending on the cell line studied. ERK1/2 signalling has a major role in ER- MDA-MB-231 cells, whereas Fra-1 accumulation occurs mainly through SPAK signalling in ER- BT549 cells. Fra-1 mutational analysis shows that the phosphorylation of S265, T223 and T230 is critical for PKCθ-driven Fra-1 stabilisation. Phosphorylation of the protein was confirmed using specific antisera against Fra-1 phosphorylated on T223 or S265. In addition, Fra-1 participates in PKCθ-induced cell invasion and is necessary for PKCθ-induced cell migration. In summary, we identified PKCθ signalling as an important regulator of Fra-1 accumulation in ER- breast cancer cells. Moreover, our results suggest that PKCθ could participate in progression of some breast cancers and could be a new therapeutic target.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Shaulian E, Karin M. AP-1 in cell proliferation and survival. Oncogene. 2001;20:2390–2400. - PubMed
-
- Eferl R, Wagner EF. AP-1: a double-edged sword in tumorigenesis. Nat Rev Cancer. 2003;3:859–868. - PubMed
-
- Young MR, Colburn NH. Fra-1 a target for cancer prevention or intervention. Gene. 2006;379:1–11. - PubMed
-
- Verde P, Casalino L, Talotta F, Yaniv M, Weitzman JB. Deciphering AP-1 function in tumorigenesis: fra-ternizing on target promoters. Cell Cycle. 2007;6:2633–2639. - PubMed
-
- Mechta F, Lallemand D, Pfarr CM, Yaniv M. Transformation by ras modifies AP1 composition and activity. Oncogene. 1997;14:837–847. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
