

Cataract-linked γD-crystallin mutants have weak affinity to lens chaperones α-crystallins
- PMID: 22289178
- PMCID: PMC3282170
- DOI: 10.1016/j.febslet.2012.01.019
Cataract-linked γD-crystallin mutants have weak affinity to lens chaperones α-crystallins
Abstract
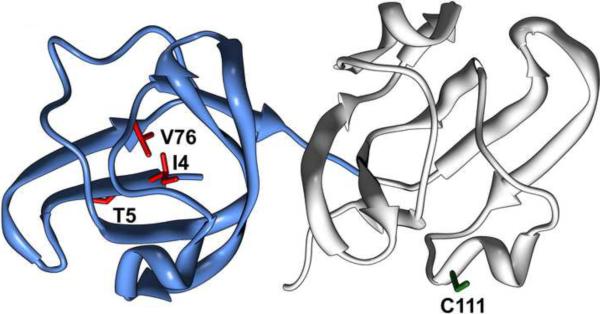
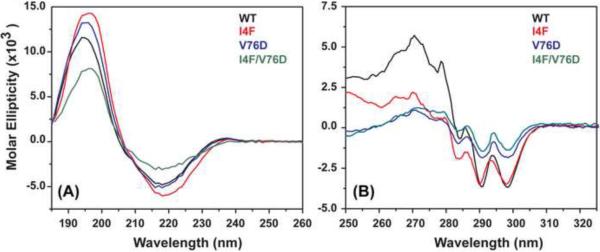
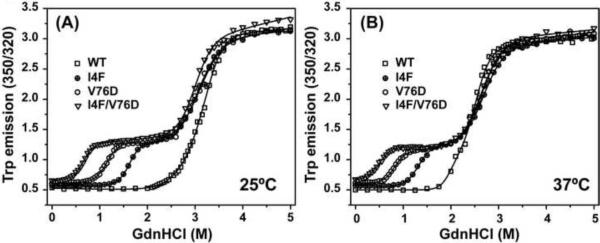
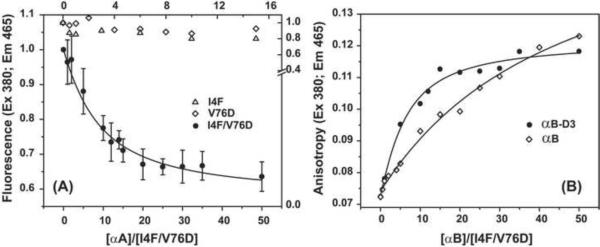
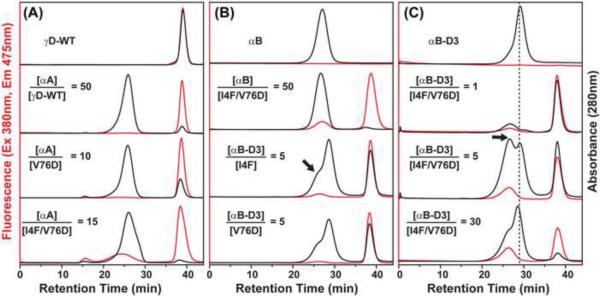
To test the hypothesis that α-crystallin chaperone activity plays a central role in maintenance of lens transparency, we investigated its interactions with γ-crystallin mutants that cause congenital cataract in mouse models. Although the two substitutions, I4F and V76D, stabilize a partially unfolded γD-crystallin intermediate, their affinities to α-crystallin are marginal even at relatively high concentrations. Detectable binding required further reduction of γD-crystallin stability which was achieved by combining the two mutations. Our results demonstrate that mutants and possibly age-damaged γ-crystallin can escape quality control by lens chaperones rationalizing the observation that they nucleate protein aggregation and lead to cataract.
Copyright © 2012 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
Figures

Similar articles
-
Cataract-causing defect of a mutant γ-crystallin proceeds through an aggregation pathway which bypasses recognition by the α-crystallin chaperone.PLoS One. 2012;7(5):e37256. doi: 10.1371/journal.pone.0037256. Epub 2012 May 24. PLoS One. 2012. PMID: 22655036 Free PMC article.
-
GammaD-crystallin associated protein aggregation and lens fiber cell denucleation.Invest Ophthalmol Vis Sci. 2007 Aug;48(8):3719-28. doi: 10.1167/iovs.06-1487. Invest Ophthalmol Vis Sci. 2007. PMID: 17652744
-
ATP differentially antagonizes the crowding-induced destabilization of human γS-crystallin and its four cataract-causing mutants.Biochem Biophys Res Commun. 2020 Dec 17;533(4):913-918. doi: 10.1016/j.bbrc.2020.09.090. Epub 2020 Sep 29. Biochem Biophys Res Commun. 2020. PMID: 33004175
-
Modulation of alpha-crystallin chaperone activity: a target to prevent or delay cataract?IUBMB Life. 2009 May;61(5):485-95. doi: 10.1002/iub.176. IUBMB Life. 2009. PMID: 19391162 Review.
-
The Functional Significance of High Cysteine Content in Eye Lens γ-Crystallins.Biomolecules. 2024 May 17;14(5):594. doi: 10.3390/biom14050594. Biomolecules. 2024. PMID: 38786000 Free PMC article. Review.
Cited by
-
α-Crystallins in the Vertebrate Eye Lens: Complex Oligomers and Molecular Chaperones.Annu Rev Phys Chem. 2021 Apr 20;72:143-163. doi: 10.1146/annurev-physchem-090419-121428. Epub 2020 Dec 15. Annu Rev Phys Chem. 2021. PMID: 33321054 Free PMC article. Review.
-
Proteinaceous Transformers: Structural and Functional Variability of Human sHsps.Int J Mol Sci. 2020 Jul 30;21(15):5448. doi: 10.3390/ijms21155448. Int J Mol Sci. 2020. PMID: 32751672 Free PMC article. Review.
-
Cataract-causing defect of a mutant γ-crystallin proceeds through an aggregation pathway which bypasses recognition by the α-crystallin chaperone.PLoS One. 2012;7(5):e37256. doi: 10.1371/journal.pone.0037256. Epub 2012 May 24. PLoS One. 2012. PMID: 22655036 Free PMC article.
-
Transgenic zebrafish models reveal distinct molecular mechanisms for cataract-linked αA-crystallin mutants.PLoS One. 2018 Nov 26;13(11):e0207540. doi: 10.1371/journal.pone.0207540. eCollection 2018. PLoS One. 2018. PMID: 30475834 Free PMC article.
-
Expression of Cataract-linked γ-Crystallin Variants in Zebrafish Reveals a Proteostasis Network That Senses Protein Stability.J Biol Chem. 2016 Dec 2;291(49):25387-25397. doi: 10.1074/jbc.M116.749606. Epub 2016 Oct 21. J Biol Chem. 2016. PMID: 27770023 Free PMC article.
References
-
- Bloemendal H, de Jong W, Jaenicke R, Lubsen NH, Slingsby C, Tardieu A. Ageing and vision: structure, stability and function of lens crystallins. Progress in Biophysics & Molecular Biology. 2004;86:407–85. - PubMed
-
- Wistow GJ, Piatigorsky J. Lens crystallins: the evolution and expression of proteins for a highly specialized tissue. Annual Review of Biochemistry. 1988;57:479–504. - PubMed
-
- Benedek GB. Theory of transparency of the eyes. Appl. Opt. 1971;10:459. - PubMed
-
- Tardieu A. α-Crystallin quaternary structure and interactive properties control eye lens transparency. International Journal of Biological Macromolecules. 1998;22:211–7. - PubMed
-
- Hanson SR, Hasan A, Smith DL, Smith JB. The major in vivo modifications of the human water-insoluble lens crystallins are disulfide bonds, deamidation, methionine oxidation and backbone cleavage. Experimental Eye Research. 2000;71:195–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
