

Deletion of TnAbaR23 results in both expected and unexpected antibiogram changes in a multidrug-resistant Acinetobacter baumannii strain
- PMID: 22290963
- PMCID: PMC3318347
- DOI: 10.1128/AAC.05334-11
Deletion of TnAbaR23 results in both expected and unexpected antibiogram changes in a multidrug-resistant Acinetobacter baumannii strain
Abstract
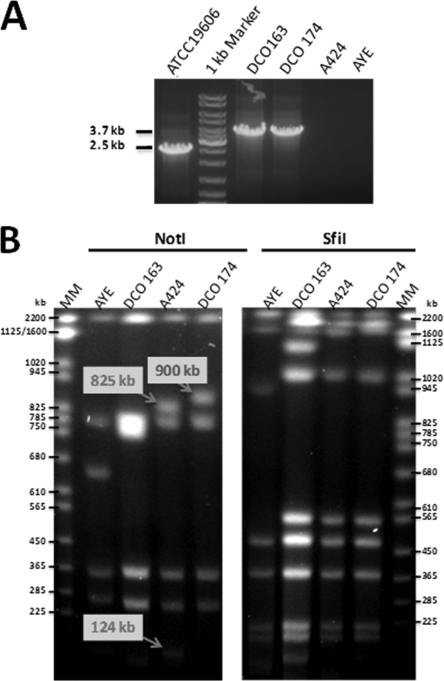
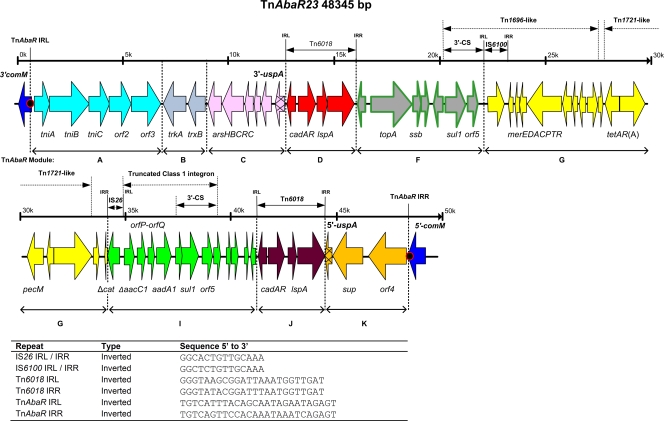
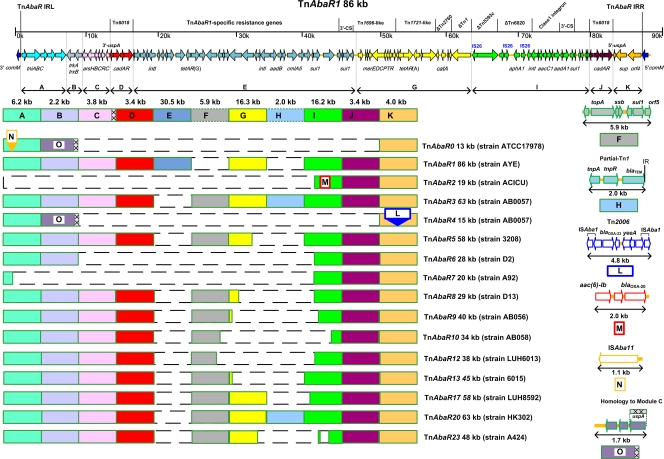
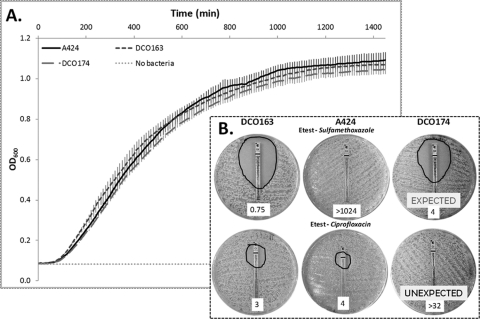
Since the 2006 discovery of the Acinetobacter baumannii strain AYE AbaR1 resistance island, similar elements have been reported in numerous members of this species. As AbaR1 is distantly related to Tn7, we have renamed it TnAbaR1. TnAbaR transposons are known to carry multiple antibiotic resistance- and efflux-associated genes, although none have been experimentally studied en bloc. We deleted the TnAbaR transposon in A. baumannii A424, which we have designated TnAbaR23, and characterized independent deletion mutants DCO163 and DCO174. The NotI pulsed-field gel electrophoresis (PFGE) profile of strain DCO174 was consistent with targeted deletion of TnAbaR23 alone, but strain DCO163 apparently harbored a second large genomic deletion. Nevertheless, "subtractive amplification" targeting 52 TnAbaR and/or resistance-associated loci yielded identical results for both mutants and highlighted genes lost relative to strain A424. PCR mapping and genome sequencing revealed the entire 48.3-kb sequence of TnAbaR23. Consistent with TnAbaR23 carrying two copies of sul1, both mutants exhibited markedly increased susceptibility to sulfamethoxazole. In contrast, loss of tetAR(A) resulted in only a minor and variable increase in tetracycline susceptibility. Despite not exhibiting a growth handicap, strain DCO163 was more susceptible than strain DCO174 to 9 of 10 antibiotics associated with mutant-to-mutant variation in susceptibility, suggesting impairment of an undefined resistance-associated function. Remarkably, despite all three strains sharing identical gyrA and parC sequences, the ciprofloxacin MIC of DCO174 was >8-fold that of DCO163 and A424, suggesting a possible paradoxical role for TnAbaR23 in promoting sensitivity to ciprofloxacin. This study highlights the importance of experimental scrutiny and challenges the assumption that resistance phenotypes can reliably be predicted from genotypes alone.
Figures
References
-
- Andrews J. 2010. BSAC methods for antimicrobial susceptibility testing. Version 9.1. March 2010 British Society for Antimicrobial Chemotherapy, Birmingham, United Kingdom: http://www.bsac.org.uk/Resources/BSAC/Version_9.1_March_2010_final.pdf
-
- Bailey JK, Pinyon JL, Anantham S, Hall RM. 2011. Distribution of the blaTEM gene and blaTEM-containing transposons in commensal Escherichia coli. J. Antimicrob. Chemother. 66:745–751 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
