

Proteome of Hydra nematocyst
- PMID: 22291027
- PMCID: PMC3323026
- DOI: 10.1074/jbc.M111.328203
Proteome of Hydra nematocyst
Abstract
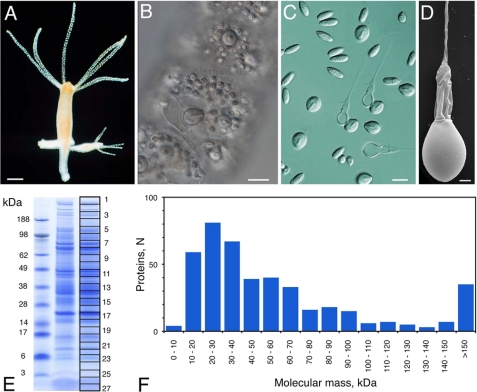
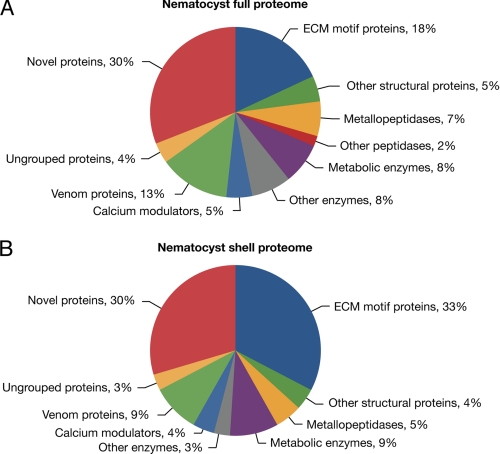
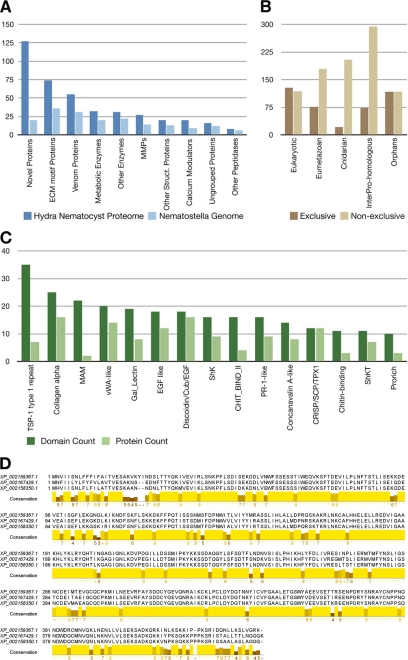
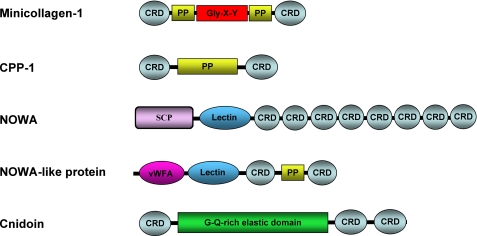
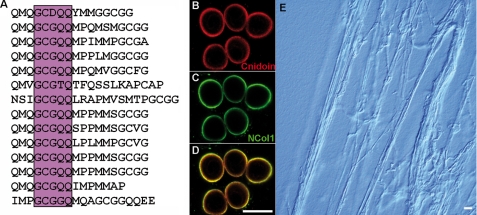
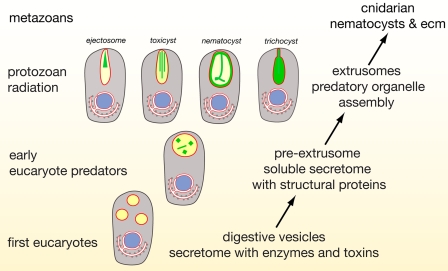
Stinging cells or nematocytes of jellyfish and other cnidarians represent one of the most poisonous and sophisticated cellular inventions in animal evolution. This ancient cell type is unique in containing a giant secretory vesicle derived from the Golgi apparatus. The organelle structure within the vesicle comprises an elastically stretched capsule (nematocyst) to which a long tubule is attached. During exocytosis, the barbed part of the tubule is accelerated with >5 million g in <700 ns, enabling a harpoon-like discharge (Nüchter, T., Benoit, M., Engel, U., Ozbek, S., and Holstein, T. W. (2006) Curr. Biol. 16, R316-R318). Hitherto, the molecular components responsible for the organelle's biomechanical properties were largely unknown. Here, we describe the proteome of nematocysts from the freshwater polyp Hydra magnipapillata. Our analysis revealed an unexpectedly complex secretome of 410 proteins with venomous and lytic but also adhesive or fibrous properties. In particular, the insoluble fraction of the nematocyst represents a functional extracellular matrix structure of collagenous and elastic nature. This finding suggests an evolutionary scenario in which exocytic vesicles harboring a venomous secretome assembled a sophisticated predatory structure from extracellular matrix motif proteins.
Figures
References
-
- Nüchter T., Benoit M., Engel U., Ozbek S., Holstein T. W. (2006) Nanosecond-scale kinetics of nematocyst discharge. Curr. Biol. 16, R316–R318 - PubMed
-
- Fry B. G., Wroe S., Teeuwisse W., van Osch M. J., Moreno K., Ingle J., McHenry C., Ferrara T., Clausen P., Scheib H., Winter K. L., Greisman L., Roelants K., van der Weerd L., Clemente C. J., Giannakis E., Hodgson W. C., Luz S., Martelli P., Krishnasamy K., Kochva E., Kwok H. F., Scanlon D., Karas J., Citron D. M., Goldstein E. J., McNaughtan J. E., Norman J. A. (2009) A central role for venom in predation by Varanus komodoensis (Komodo dragon) and the extinct giant Varanus (Megalania) priscus. Proc. Natl. Acad. Sci. U.S.A. 106, 8969–8974 - PMC - PubMed
-
- Fry B. G., Roelants K., Champagne D. E., Scheib H., Tyndall J. D., King G. F., Nevalainen T. J., Norman J. A., Lewis R. J., Norton R. S., Renjifo C., de la Vega R. C. (2009) The toxicogenomic multiverse: convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 10, 483–511 - PubMed
-
- Eble J. A. (2010) Matrix biology meets toxinology. Matrix Biol. 29, 239–247 - PubMed
-
- Mariscal R. N. (1974) Scanning electron microscopy of the sensory surface of the tentacles of sea anemones and corals. Z. Zellforsch. 147, 149–156 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
