

Loss of function of FATTY ACID DESATURASE7 in tomato enhances basal aphid resistance in a salicylate-dependent manner
- PMID: 22291202
- PMCID: PMC3320204
- DOI: 10.1104/pp.111.191262
Loss of function of FATTY ACID DESATURASE7 in tomato enhances basal aphid resistance in a salicylate-dependent manner
Abstract
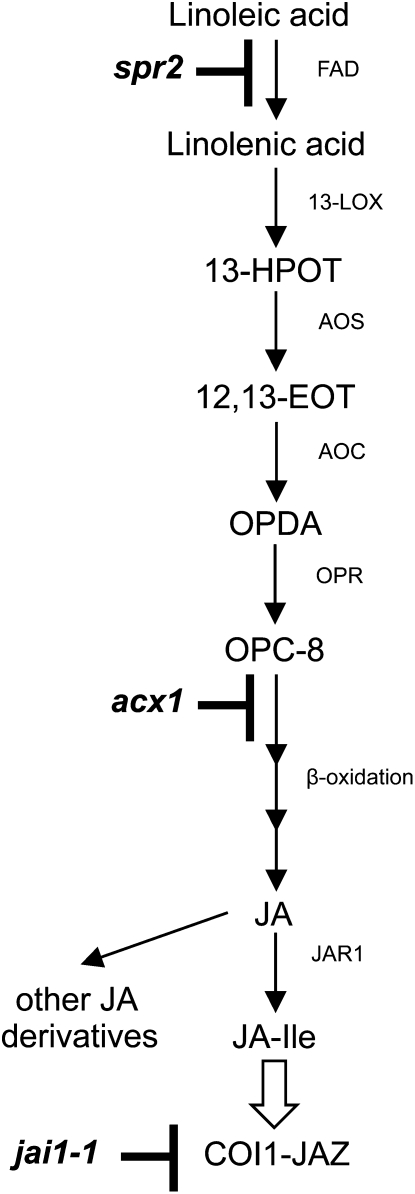
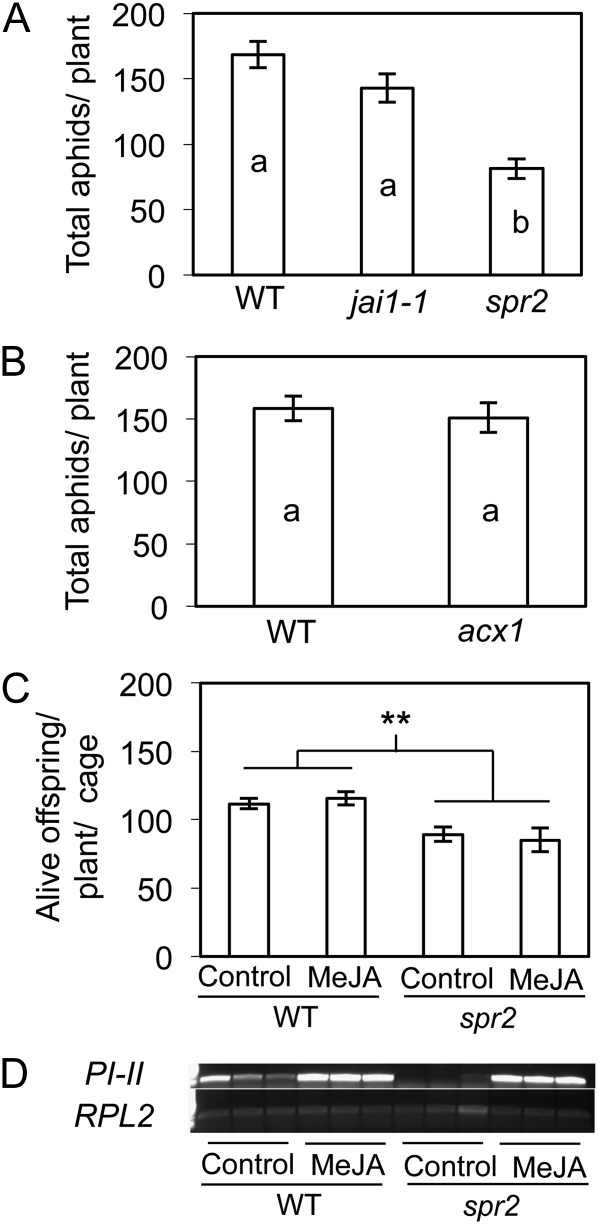
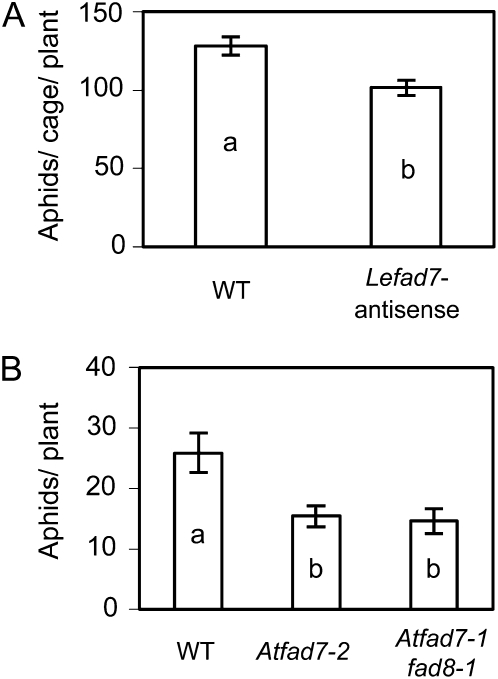
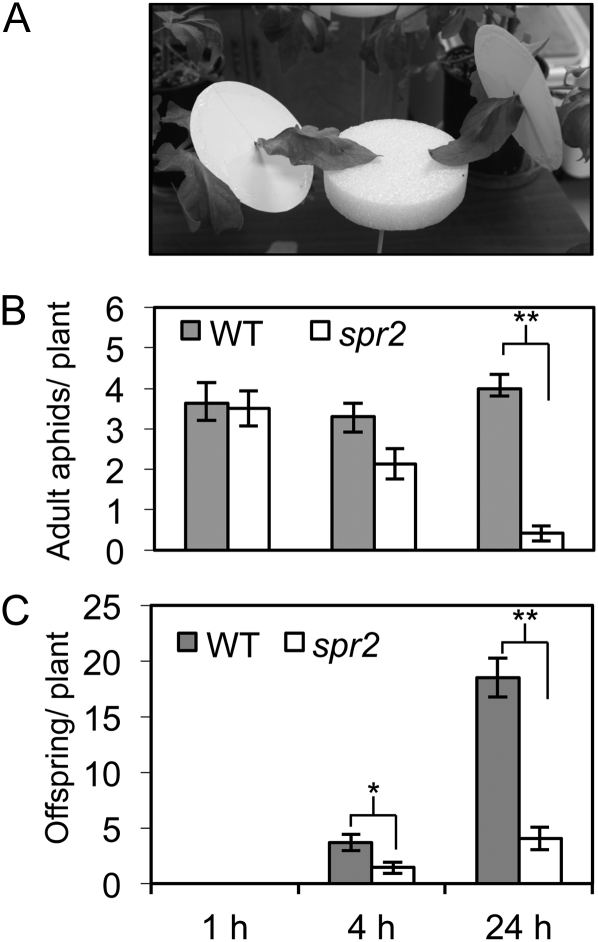
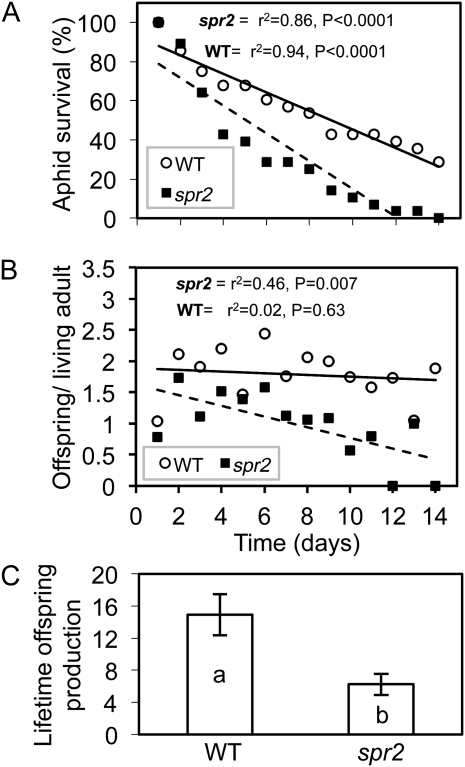
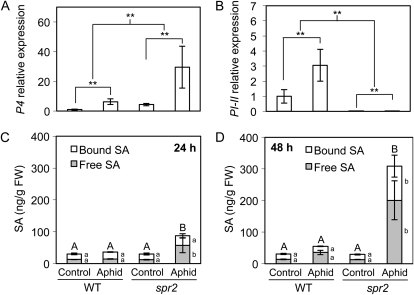
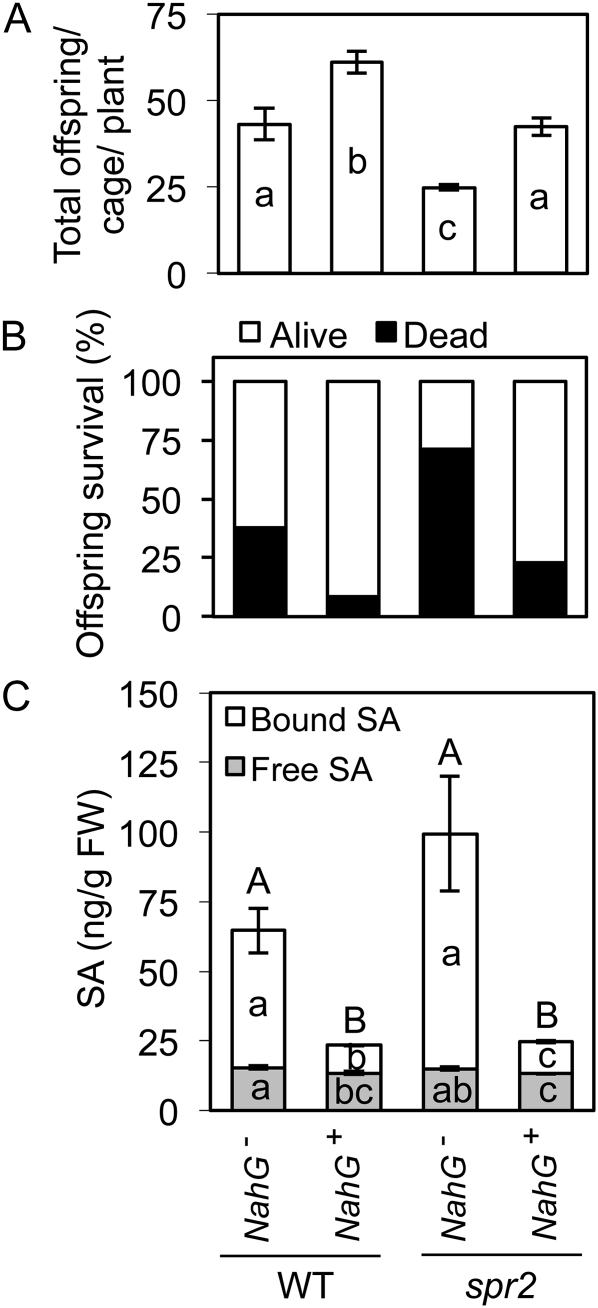
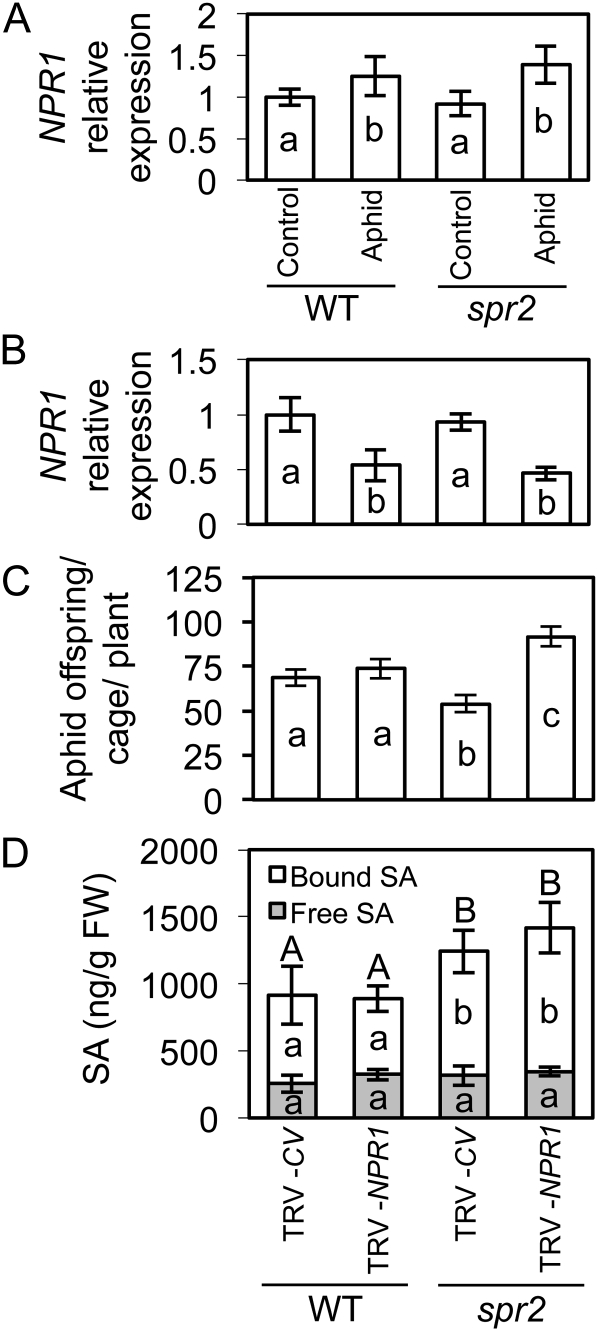
We report here that disruption of function of the ω-3 FATTY ACID DESATURASE7 (FAD7) enhances plant defenses against aphids. The suppressor of prosystemin-mediated responses2 (spr2) mutation in tomato (Solanum lycopersicum), which eliminates the function of FAD7, reduces the settling behavior, survival, and fecundity of the potato aphid (Macrosiphum euphorbiae). Likewise, the antisense suppression of LeFAD7 expression in wild-type tomato plants reduces aphid infestations. Aphid resistance in the spr2 mutant is associated with enhanced levels of salicylic acid (SA) and mRNA encoding the pathogenesis-related protein P4. Introduction of the Naphthalene/salicylate hydroxylase transgene, which suppresses SA accumulation, restores wild-type levels of aphid susceptibility to spr2. Resistance in spr2 is also lost when we utilize virus-induced gene silencing to suppress the expression of NONEXPRESSOR OF PATHOGENESIS-RELATED PROTEINS1 (NPR1), a positive regulator of many SA-dependent defenses. These results indicate that FAD7 suppresses defenses against aphids that are mediated through SA and NPR1. Although loss of function of FAD7 also inhibits the synthesis of jasmonate (JA), the effects of this desaturase on aphid resistance are not dependent on JA; other mutants impaired in JA synthesis (acx1) or perception (jai1-1) show wild-type levels of aphid susceptibility, and spr2 retains aphid resistance when treated with methyl jasmonate. Thus, FAD7 may influence JA-dependent defenses against chewing insects and SA-dependent defenses against aphids through independent effects on JA synthesis and SA signaling. The Arabidopsis (Arabidopsis thaliana) mutants Atfad7-2 and Atfad7-1fad8 also show enhanced resistance to the green peach aphid (Myzus persicae) compared with wild-type controls, indicating that FAD7 influences plant-aphid interactions in at least two plant families.
Figures
References
-
- Berberich T, Harada M, Sugawara K, Kodama H, Iba K, Kusano T. (1998) Two maize genes encoding ω-3 fatty acid desaturase and their differential expression to temperature. Plant Mol Biol 36: 297–306 - PubMed
-
- Bhattarai KK, Xie Q-G, Pourshalimi D, Younglove T, Kaloshian I. (2007) Coil-dependent signaling pathway is not required for Mi-1-mediated potato aphid resistance. Mol Plant Microbe Interact 20: 276–282 - PubMed
-
- Blée E. (2002) Impact of phyto-oxylipins in plant defense. Trends Plant Sci 7: 315–322 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
