

The chromosomal passenger complex activates Polo kinase at centromeres
- PMID: 22291575
- PMCID: PMC3265468
- DOI: 10.1371/journal.pbio.1001250
The chromosomal passenger complex activates Polo kinase at centromeres
Erratum in
- PLoS Biol. 2012 Feb;10(2). doi:10.1371/annotation/7832f573-e0d9-465f-b5d4-0ac1014b6112
Abstract
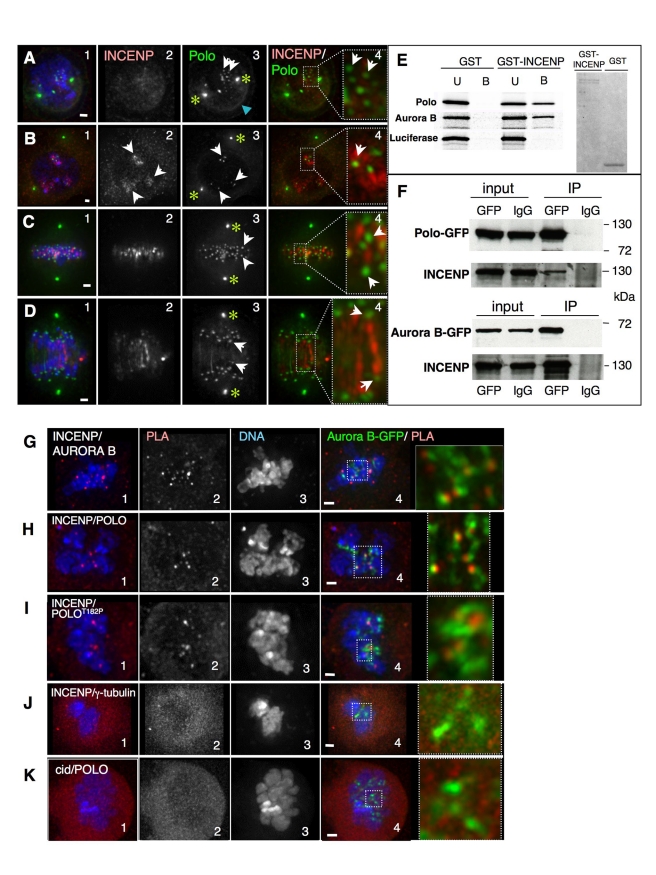
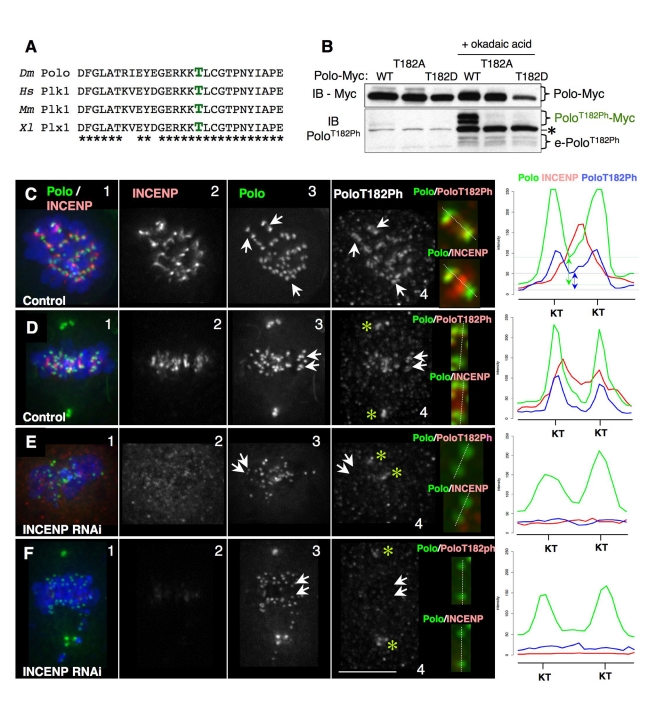
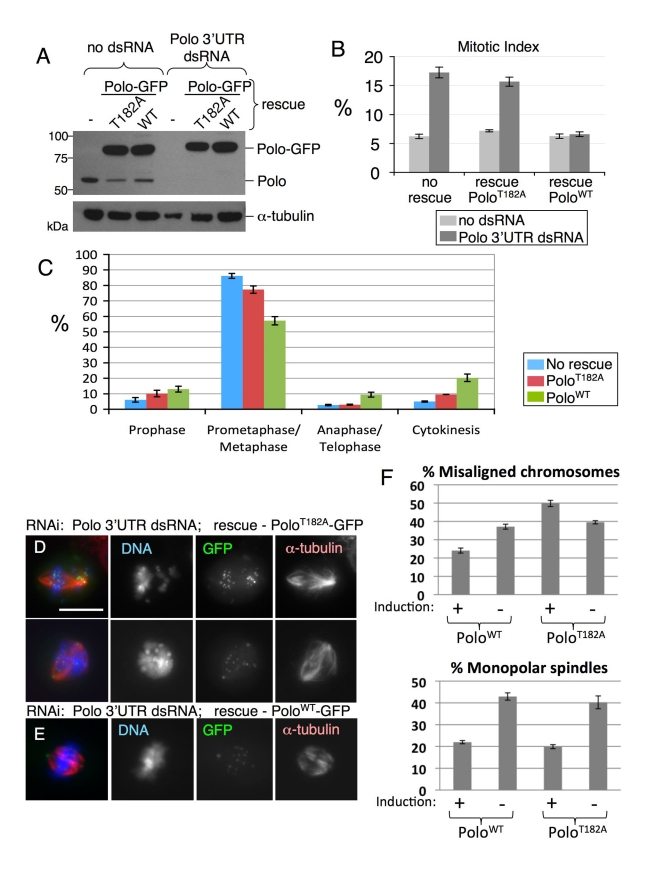
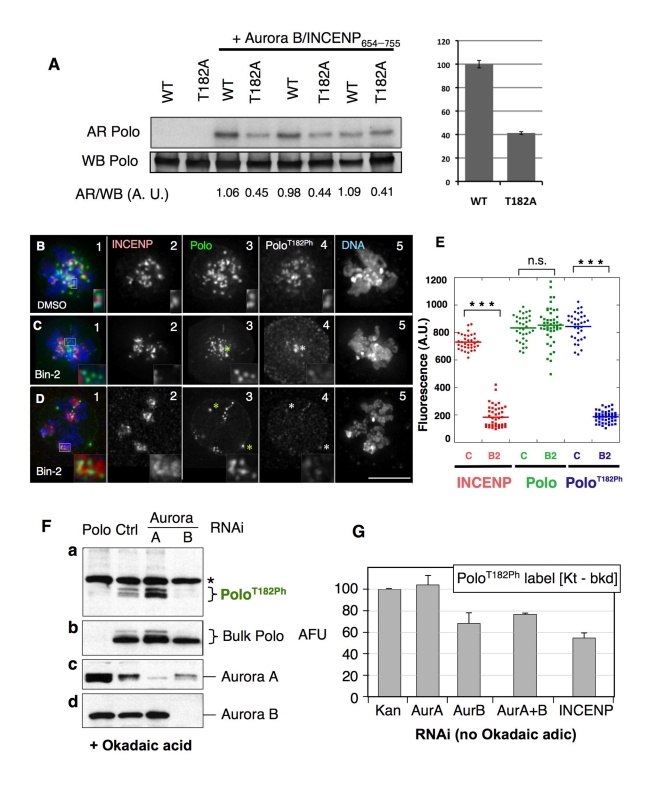
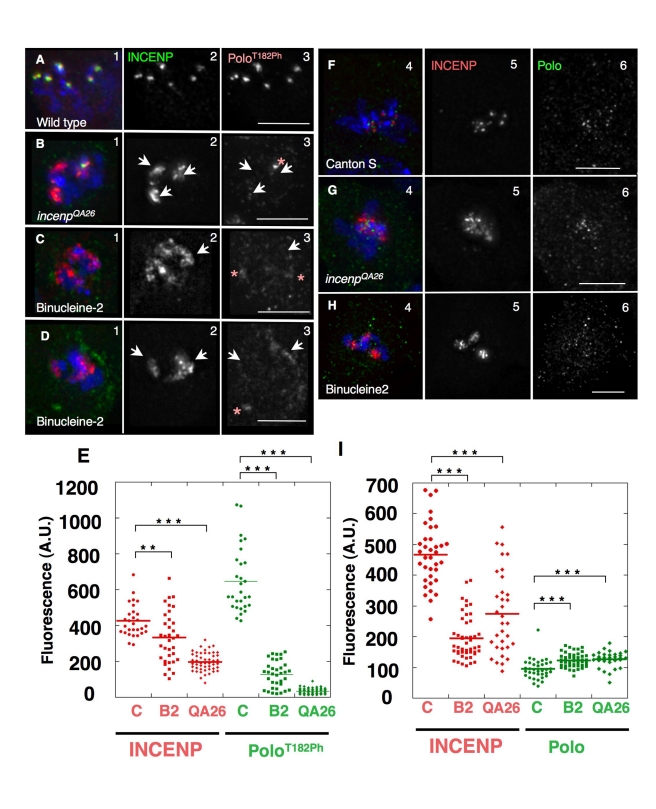
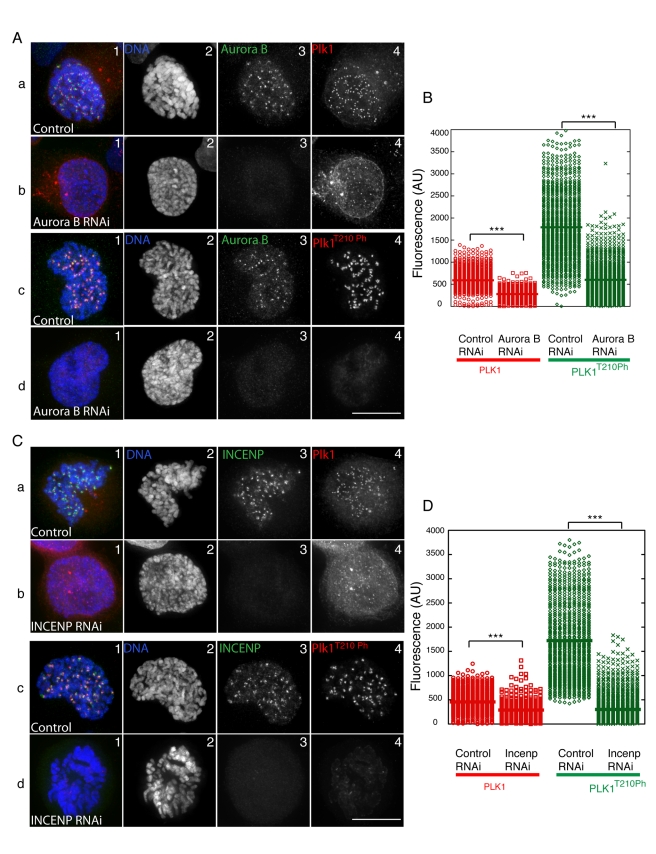
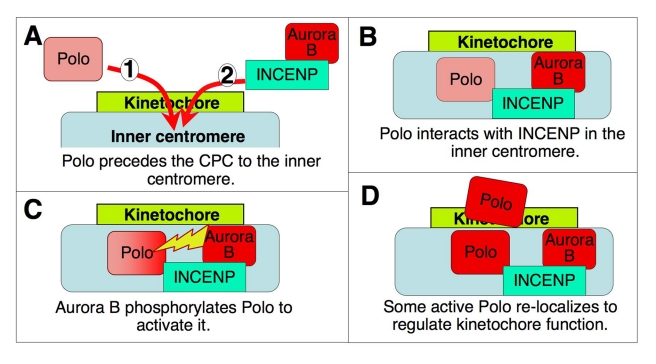
The coordinated activities at centromeres of two key cell cycle kinases, Polo and Aurora B, are critical for ensuring that the two sister kinetochores of each chromosome are attached to microtubules from opposite spindle poles prior to chromosome segregation at anaphase. Initial attachments of chromosomes to the spindle involve random interactions between kinetochores and dynamic microtubules, and errors occur frequently during early stages of the process. The balance between microtubule binding and error correction (e.g., release of bound microtubules) requires the activities of Polo and Aurora B kinases, with Polo promoting stable attachments and Aurora B promoting detachment. Our study concerns the coordination of the activities of these two kinases in vivo. We show that INCENP, a key scaffolding subunit of the chromosomal passenger complex (CPC), which consists of Aurora B kinase, INCENP, Survivin, and Borealin/Dasra B, also interacts with Polo kinase in Drosophila cells. It was known that Aurora A/Bora activates Polo at centrosomes during late G2. However, the kinase that activates Polo on chromosomes for its critical functions at kinetochores was not known. We show here that Aurora B kinase phosphorylates Polo on its activation loop at the centromere in early mitosis. This phosphorylation requires both INCENP and Aurora B activity (but not Aurora A activity) and is critical for Polo function at kinetochores. Our results demonstrate clearly that Polo kinase is regulated differently at centrosomes and centromeres and suggest that INCENP acts as a platform for kinase crosstalk at the centromere. This crosstalk may enable Polo and Aurora B to achieve a balance wherein microtubule mis-attachments are corrected, but proper attachments are stabilized allowing proper chromosome segregation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- van Vugt M. A, Medema R. H. Getting in and out of mitosis with Polo-like kinase-1. Oncogene. 2005;24:2844–2859. - PubMed
-
- Petronczki M, Lenart P, Peters J. M. Polo on the rise—from mitotic entry to cytokinesis with Plk1. Dev Cell. 2008;14:646–659. - PubMed
-
- Archambault V, Glover D. M. Polo-like kinases: conservation and divergence in their functions and regulation. Nat Rev Mol Cell Biol. 2009;10:265–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
