

Methanogenic pathway and archaeal communities in three different anoxic soils amended with rice straw and maize straw
- PMID: 22291691
- PMCID: PMC3265783
- DOI: 10.3389/fmicb.2012.00004
Methanogenic pathway and archaeal communities in three different anoxic soils amended with rice straw and maize straw
Abstract
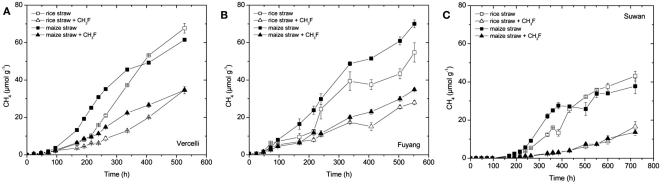
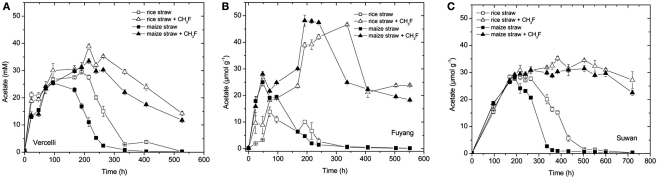
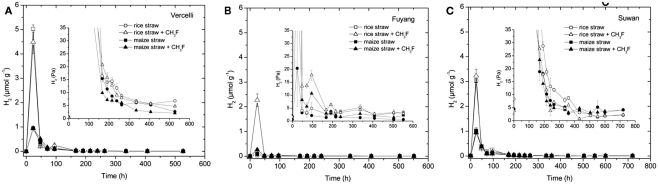
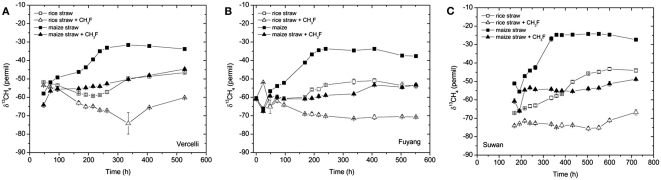
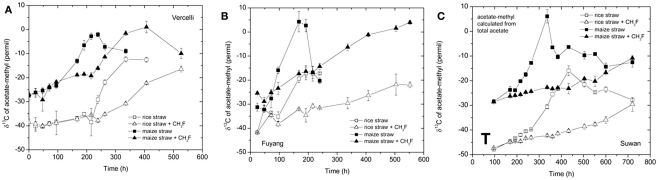
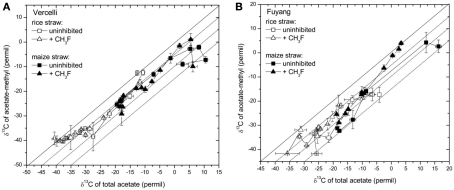
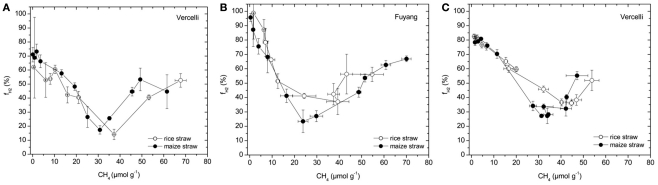
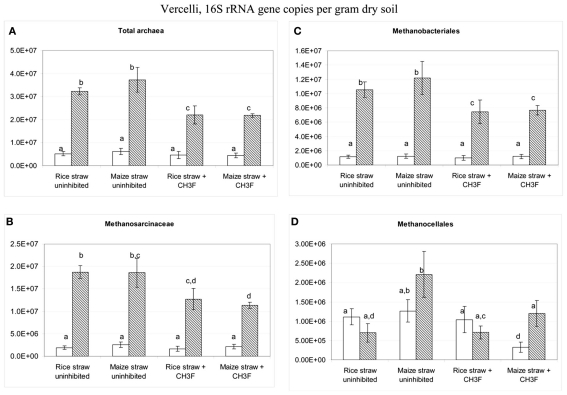
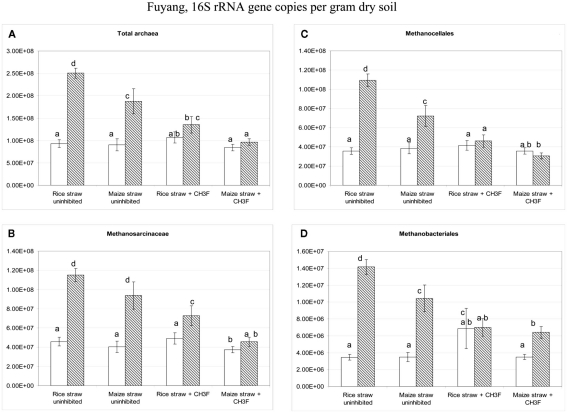
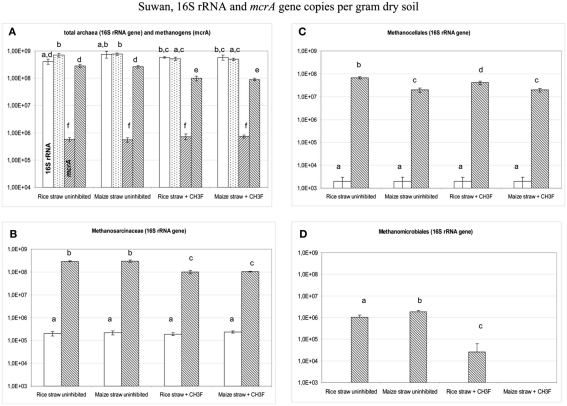
Addition of straw is common practice in rice agriculture, but its effect on the path of microbial CH(4) production and the microbial community involved is not well known. Since straw from rice (C3 plant) and maize plants (C4 plant) exhibit different δ(13)C values, we compared the effect of these straw types using anoxic rice field soils from Italy and China, and also a soil from Thailand that had previously not been flooded. The temporal patterns of production of CH(4) and its major substrates H(2) and acetate, were slightly different between rice straw and maize straw. Addition of methyl fluoride, an inhibitor of acetoclastic methanogenesis, resulted in partial inhibition of acetate consumption and CH(4) production. The δ(13)C of the accumulated CH(4) and acetate reflected the different δ(13)C values of rice straw versus maize straw. However, the relative contribution of hydrogenotrophic methanogenesis to total CH(4) production exhibited a similar temporal change when scaled to CH(4) production irrespectively of whether rice straw or maize straw was applied. The composition of the methanogenic archaeal communities was characterized by terminal restriction fragment length polymorphism (T-RFLP) analysis and was quantified by quantitative PCR targeting archaeal 16S rRNA genes or methanogenic mcrA genes. The size of the methanogenic communities generally increased during incubation with straw, but the straw type had little effect. Instead, differences were found between the soils, with Methanosarcinaceae and Methanobacteriales dominating straw decomposition in Italian soil, Methanosarcinaceae, Methanocellales, and Methanobacteriale in China soil, and Methanosarcinaceae and Methanocellales in Thailand soil. The experiments showed that methanogenic degradation in different soils involved different methanogenic population dynamics. However, the path of CH(4) production was hardly different between degradation of rice straw versus maize straw and was also similar for the different soil types.
Keywords: archaea; methanogenesis; pathway; rice field soil; straw.
Figures
References
-
- Amelung W., Brodowski S., Sandhage-Hofmann A., Bol R. (2008). Combining biomarker with stable isotope analyses for assessing the transformation and turnover of soil organic matter. Adv. Agron. 100, 155–250 10.1016/S0065-2113(08)00606-8 - DOI
-
- Balesdent J., Balabane M. (1992). Maize root-derived soil organic carbon estimated by natural C-13 abundance. Soil Biol. Biochem. 24, 97–101 10.1016/0038-0717(92)90264-X - DOI
-
- Bilek R. S., Tyler S. C., Sass R. L., Fisher F. M. (1999). Differences in CH4 oxidation and pathways of production between rice cultivars deduced from measurements of CH4 flux and δ13C of CH4 and CO2. Global Biogeochem. Cycles 13, 1029–1044 10.1029/1999GB900040 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous