

Hymyc1 downregulation promotes stem cell proliferation in Hydra vulgaris
- PMID: 22292012
- PMCID: PMC3264606
- DOI: 10.1371/journal.pone.0030660
Hymyc1 downregulation promotes stem cell proliferation in Hydra vulgaris
Abstract
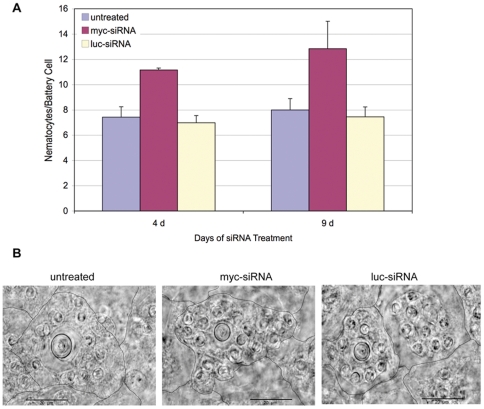
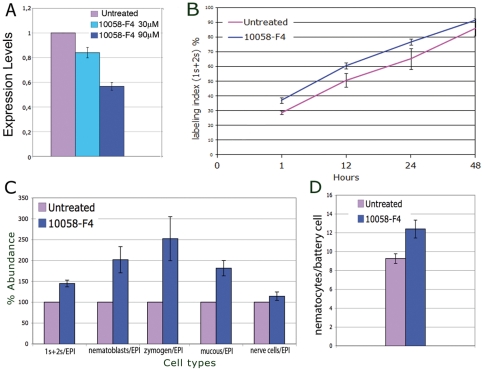
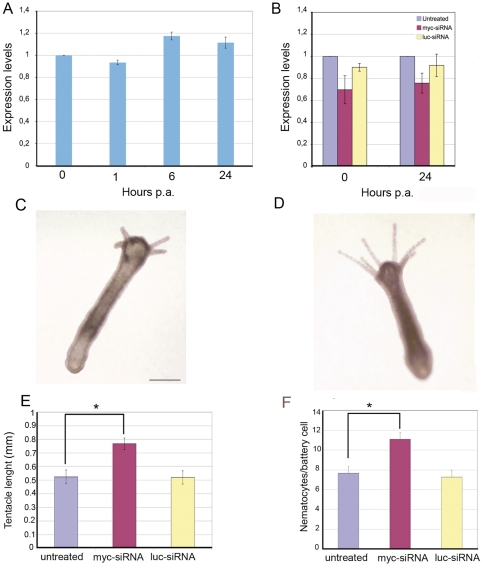
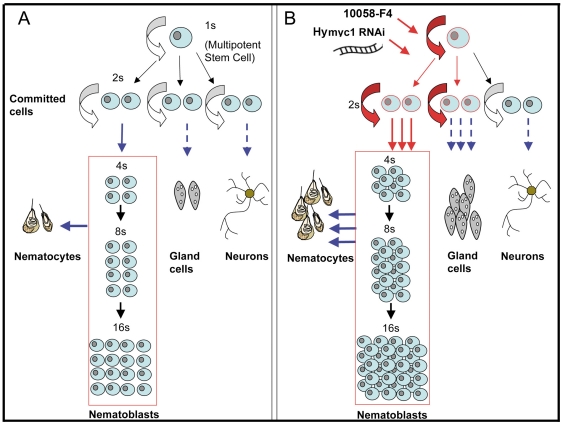
Hydra is a unique model for studying the mechanisms underlying stem cell biology. The activity of the three stem cell lineages structuring its body constantly replenishes mature cells lost due to normal tissue turnover. By a poorly understood mechanism, stem cells are maintained through self-renewal while concomitantly producing differentiated progeny. In vertebrates, one of many genes that participate in regulating stem cell homeostasis is the protooncogene c-myc, which has been recently identified also in Hydra, and found expressed in the interstitial stem cell lineage. In the present paper, by developing a novel strategy of RNA interference-mediated gene silencing (RNAi) based on an enhanced uptake of small interfering RNAi (siRNA), we provide molecular and biological evidence for an unexpected function of the Hydra myc gene (Hymyc1) in the homeostasis of the interstitial stem cell lineage. We found that Hymyc1 inhibition impairs the balance between stem cell self renewal/differentiation, as shown by the accumulation of stem cell intermediate and terminal differentiation products in genetically interfered animals. The identical phenotype induced by the 10058-F4 inhibitor, a disruptor of c-Myc/Max dimerization, demonstrates the specificity of the RNAi approach. We show the kinetic and the reversible feature of Hymyc1 RNAi, together with the effects displayed on regenerating animals. Our results show the involvement of Hymyc1 in the control of interstitial stem cell dynamics, provide new clues to decipher the molecular control of the cell and tissue plasticity in Hydra, and also provide further insights into the complex myc network in higher organisms. The ability of Hydra cells to uptake double stranded RNA and to trigger a RNAi response lays the foundations of a comprehensive analysis of the RNAi response in Hydra allowing us to track back in the evolution and the origin of this process.
Conflict of interest statement
Figures
Similar articles
-
Stem cell-specific activation of an ancestral myc protooncogene with conserved basic functions in the early metazoan Hydra.Proc Natl Acad Sci U S A. 2010 Mar 2;107(9):4051-6. doi: 10.1073/pnas.0911060107. Epub 2010 Feb 8. Proc Natl Acad Sci U S A. 2010. PMID: 20142507 Free PMC article.
-
DNA-Coated Gold Nanoparticles for the Detection of mRNA in Live Hydra Vulgaris Animals.ACS Appl Mater Interfaces. 2019 Apr 17;11(15):13905-13911. doi: 10.1021/acsami.8b17846. Epub 2018 Dec 11. ACS Appl Mater Interfaces. 2019. PMID: 30525369
-
Deficient autophagy in epithelial stem cells drives aging in the freshwater cnidarian Hydra.Development. 2020 Jan 23;147(2):dev177840. doi: 10.1242/dev.177840. Development. 2020. PMID: 31862842 Free PMC article.
-
The Hydra polyp: nothing but an active stem cell community.Dev Growth Differ. 2010 Jan;52(1):15-25. doi: 10.1111/j.1440-169X.2009.01143.x. Epub 2009 Nov 5. Dev Growth Differ. 2010. PMID: 19891641 Review.
-
Stemness in Hydra - a current perspective.Int J Dev Biol. 2012;56(6-8):509-17. doi: 10.1387/ijdb.113426bh. Int J Dev Biol. 2012. PMID: 22689357 Review.
Cited by
-
Molecular signature of an ancient organizer regulated by Wnt/β-catenin signalling during primary body axis patterning in Hydra.Commun Biol. 2019 Nov 26;2:434. doi: 10.1038/s42003-019-0680-3. eCollection 2019. Commun Biol. 2019. PMID: 31799436 Free PMC article.
-
Hydra myc2, a unique pre-bilaterian member of the myc gene family, is activated in cell proliferation and gametogenesis.Biol Open. 2014 Apr 25;3(5):397-407. doi: 10.1242/bio.20147005. Biol Open. 2014. PMID: 24771621 Free PMC article.
-
Stem cell dynamics in Cnidaria: are there unifying principles?Dev Genes Evol. 2013 Mar;223(1-2):53-66. doi: 10.1007/s00427-012-0429-1. Epub 2012 Nov 21. Dev Genes Evol. 2013. PMID: 23179637 Free PMC article. Review.
-
Multi-functionality and plasticity characterize epithelial cells in Hydra.Tissue Barriers. 2015 Jul 15;3(4):e1068908. doi: 10.1080/21688370.2015.1068908. eCollection 2015 Oct-Dec. Tissue Barriers. 2015. PMID: 26716072 Free PMC article. Review.
-
Developmental Progression in the Coral Acropora digitifera Is Controlled by Differential Expression of Distinct Regulatory Gene Networks.Genome Biol Evol. 2016 Mar 23;8(3):851-70. doi: 10.1093/gbe/evw042. Genome Biol Evol. 2016. PMID: 26941230 Free PMC article.
References
-
- Askew DS, Ashmun RA, Simmons BC, Cleveland JL. Constitutive c-myc expression in an IL-3-dependent myeloid cell line suppresses cell cycle arrest and accelerates apoptosis. Oncogene. 1991;6:1915–1922. - PubMed
-
- Bosch TC. Hydra and the evolution of stem cells. Bioessays. 2009;31:478–486. - PubMed
-
- Bode HR. The interstitial cell lineage of hydra: a stem cell system that arose early in evolution. J Cell Sci. 1996;109(Pt 6):1155–1164. - PubMed
-
- Watanabe H, Hoang VT, Mattner R, Holstein TW. Immortality and the base of multicellular life: Lessons from cnidarian stem cells. Semin Cell Dev Biol. 2009;20:1114–1125. - PubMed
-
- Campbell RD, David CN. Cell cycle kinetics and development of Hydra attenuata. II. Interstitial cells. J Cell Sci. 1974;16:349–358. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
