

c-Ets1 inhibits the interaction of NF-κB and CREB, and downregulates IL-1β-induced MUC5AC overproduction during airway inflammation
- PMID: 22294049
- PMCID: PMC3282431
- DOI: 10.1038/mi.2011.67
c-Ets1 inhibits the interaction of NF-κB and CREB, and downregulates IL-1β-induced MUC5AC overproduction during airway inflammation
Abstract
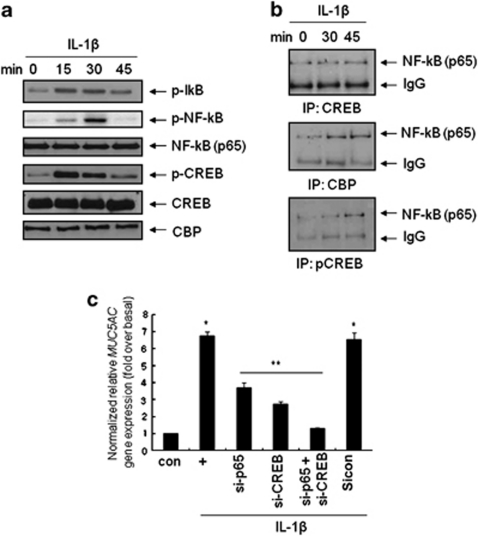
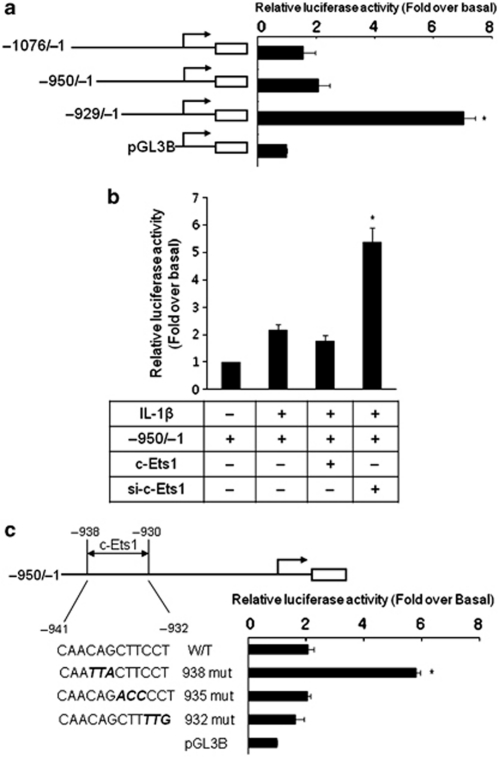
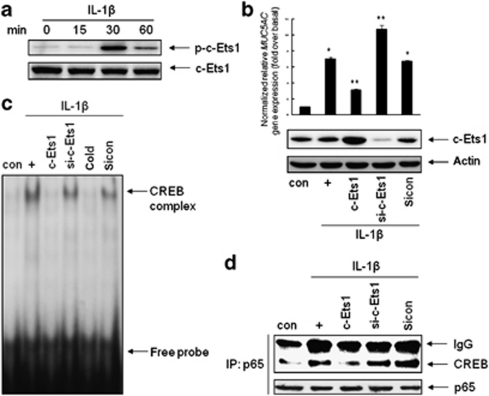
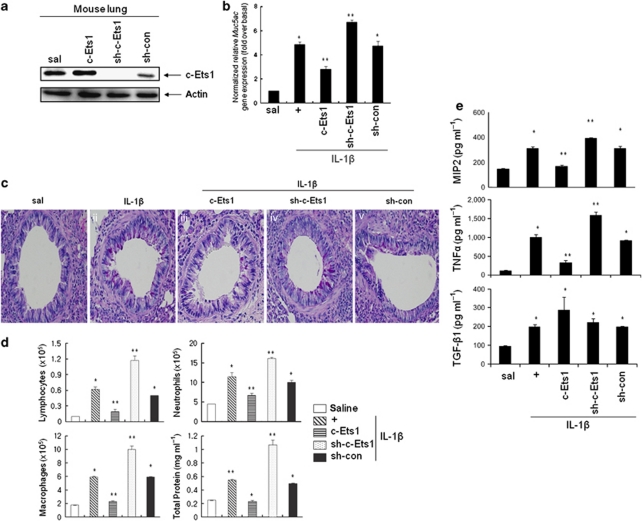
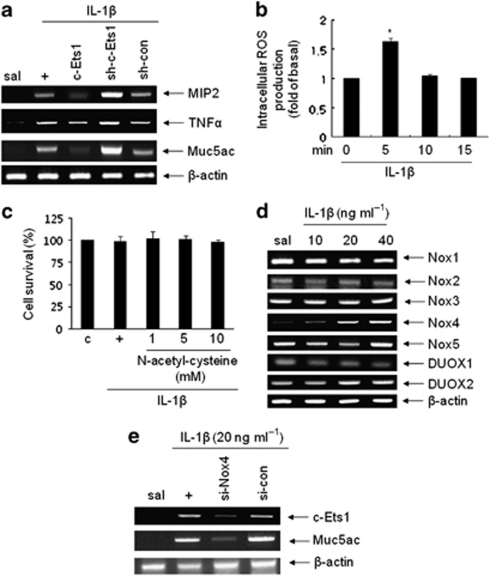
Mucin hypersecretion is frequently observed in many inflammatory diseases of the human respiratory tract. As mucin hypersecretion refers to uncontrolled mucin expression and secretion during inflammation, studies examining the negative control mechanisms of mucin hypersecretion are vital in developing novel therapeutic medications. We hypothesized that the c-Ets1 induced by interleukin (IL)-1β would decrease MUC5AC overproduction by inhibiting the interaction of NF-κB with cAMP response element-binding protein (CREB) in vivo. Stimulation with IL-1β caused the direct binding of NF-κB and CREB to the MUC5AC promoter, thus increasing MUC5AC gene expression. However, IL-1β-induced MUC5AC messenger RNA levels were surprizingly downregulated by c-Ets1 (located -938 to -930). Interestingly, c-Ets1 also suppressed IL-1β-induced MUC5AC gene expression in vitro and in vivo by disrupting the interaction of NF-κB with CREB on the MUC5AC promoter. In addition, c-Ets1 also inhibited significant morphologic changes and inflammatory cell infiltration after IL-1β exposure in mouse lungs infected with either wild-type or shRNA-c-Ets1. Moreover, reactive oxygen species produced by NOX4 increased c-Ets1 gene expression and MUC5AC gene expression in alveolar macrophages from bronchoalveolar lavage fluid. These results suggest a molecular paradigm for the establishment of a novel mechanism underlying the negative regulation of mucin overproduction, thus enhancing our understanding of airway inflammation.
Figures
References
-
- Song K.S., Kim H.J., Lee J.G. Rgulator of G-protein signaling 4 suppresses LPS-induced MUC5AC overproduction in the airway. Am. J. Respir. Cell Mol. Biol. 2009;41,:40–49. - PubMed
-
- Song K.S., Choi Y.H., Kim J.M., Lee H., Lee T.J., Yoon J.H. Suppression of prostaglandin E2-induced MUC5AC overproduction by RGS4 in the airway. Am. J. Physiol. Lung Cell Mol. Physiol. 2009;296,:L684–L692. - PubMed
-
- Song K.S., Kim K., Chung K.C., Seol J.H., Yoon J.H. Interaction of SOCS3 with NonO attenuates IL-1beta-dependent MUC8 gene expression. Biochem. Biophys. Res. Commun. 2008;377,:946–951. - PubMed
-
- Song K.S., et al. Interleukin-1 beta and tumor necrosis factor-alpha induce MUC5AC overexpression through a mechanism involving ERK/p38 mitogen-activated protein kinases-MSK1-CREB activation in human airway epithelial cells. J. Biol. Chem. 2003;278,:23243–23250. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
