

Interleukin-22 is produced by invariant natural killer T lymphocytes during influenza A virus infection: potential role in protection against lung epithelial damages
- PMID: 22294696
- PMCID: PMC3308738
- DOI: 10.1074/jbc.M111.304758
Interleukin-22 is produced by invariant natural killer T lymphocytes during influenza A virus infection: potential role in protection against lung epithelial damages
Abstract
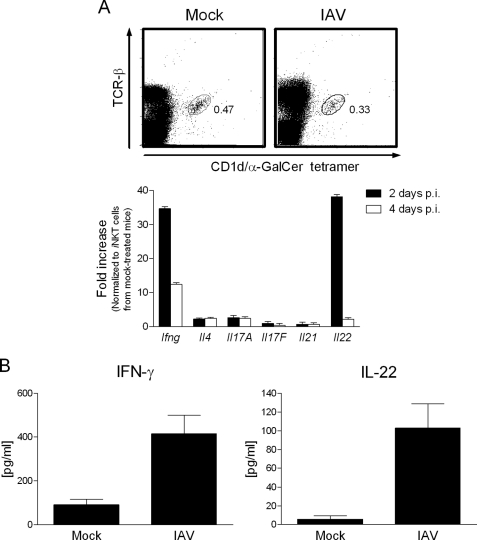
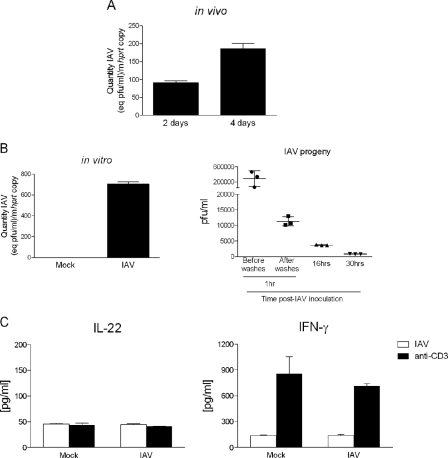
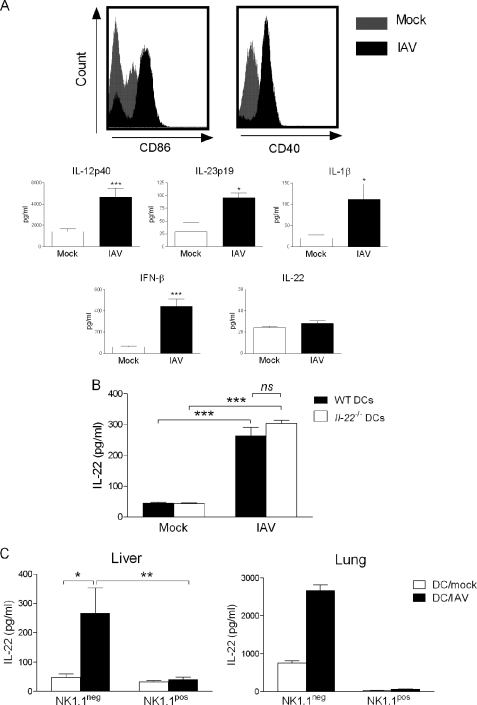
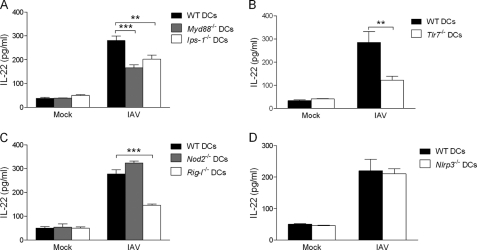
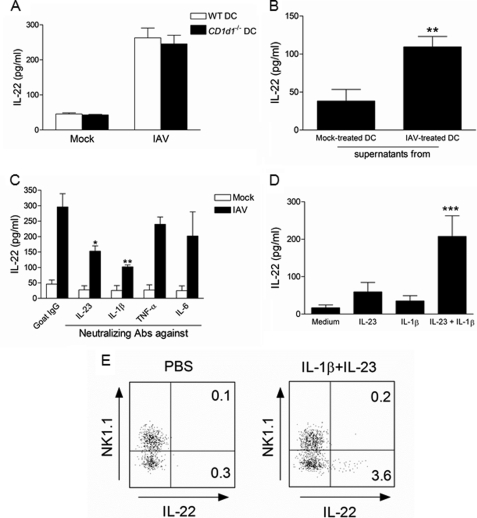
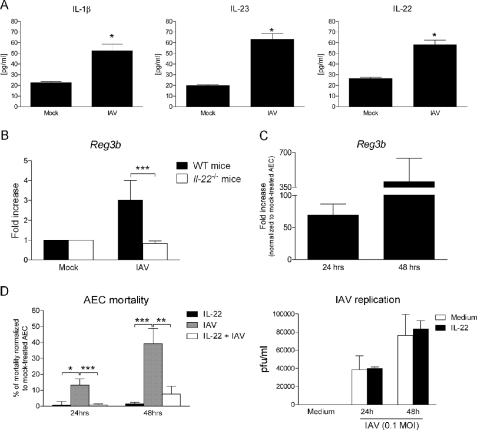
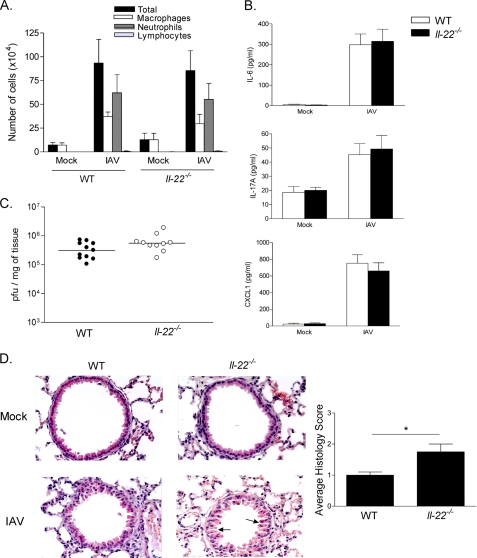
Invariant natural killer T (iNKT) cells are non-conventional lipid-reactive αβ T lymphocytes that play a key role in host responses during viral infections, in particular through the swift production of cytokines. Their beneficial role during experimental influenza A virus (IAV) infection has recently been proposed, although the mechanisms involved remain elusive. Here we show that during in vivo IAV infection, mouse pulmonary iNKT cells produce IFN-γ and IL-22, a Th17-related cytokine critical in mucosal immunity. Although permissive to viral replication, IL-22 production by iNKT cells is not due to IAV infection per se of these cells but is indirectly mediated by IAV-infected dendritic cells (DCs). We show that activation of the viral RNA sensors TLR7 and RIG-I in DCs is important for triggering IL-22 secretion by iNKT cells, whereas the NOD-like receptors NOD2 and NLRP3 are dispensable. Invariant NKT cells respond to IL-1β and IL-23 provided by infected DCs independently of the CD1d molecule to release IL-22. In vitro, IL-22 protects IAV-infected airway epithelial cells against mortality but has no role on viral replication. Finally, during early IAV infection, IL-22 plays a positive role in the control of lung epithelial damages. Overall, IAV infection of DCs activates iNKT cells, providing a rapid source of IL-22 that might be beneficial to preserve the lung epithelium integrity.
Figures
References
-
- Bendelac A., Savage P. B., Teyton L. (2007) The biology of NKT cells. Annu. Rev. Immunol. 25, 297–336 - PubMed
-
- Kronenberg M. (2005) Toward an understanding of NKT cell biology. Progress and paradoxes. Annu. Rev. Immunol. 23, 877–900 - PubMed
-
- Van Kaer L., Joyce S. (2005) Innate immunity. NKT cells in the spotlight. Curr. Biol. 15, R429–R431 - PubMed
-
- Kawano T., Cui J., Koezuka Y., Toura I., Kaneko Y., Motoki K., Ueno H., Nakagawa R., Sato H., Kondo E., Koseki H., Taniguchi M. (1997) CD1d-restricted and TCR-mediated activation of vα14 NKT cells by glycosylceramides. Science 278, 1626–1629 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
