

Two tyrosyl radicals stabilize high oxidation states in cytochrome C oxidase for efficient energy conservation and proton translocation
- PMID: 22296274
- PMCID: PMC3418888
- DOI: 10.1021/ja210535w
Two tyrosyl radicals stabilize high oxidation states in cytochrome C oxidase for efficient energy conservation and proton translocation
Abstract
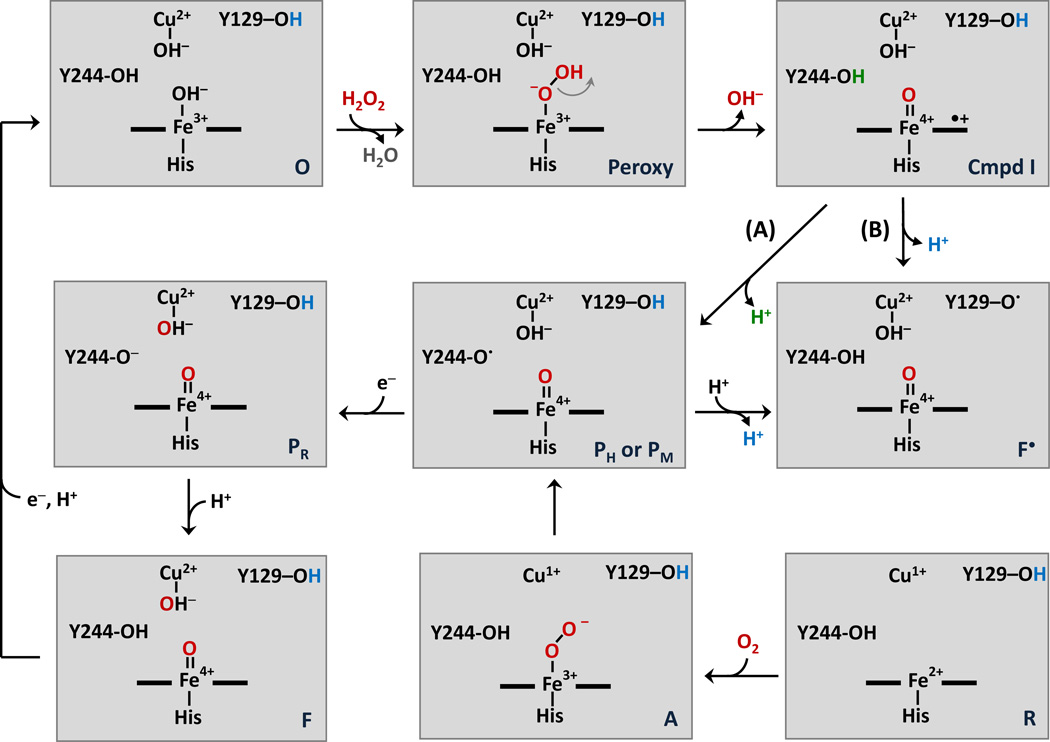
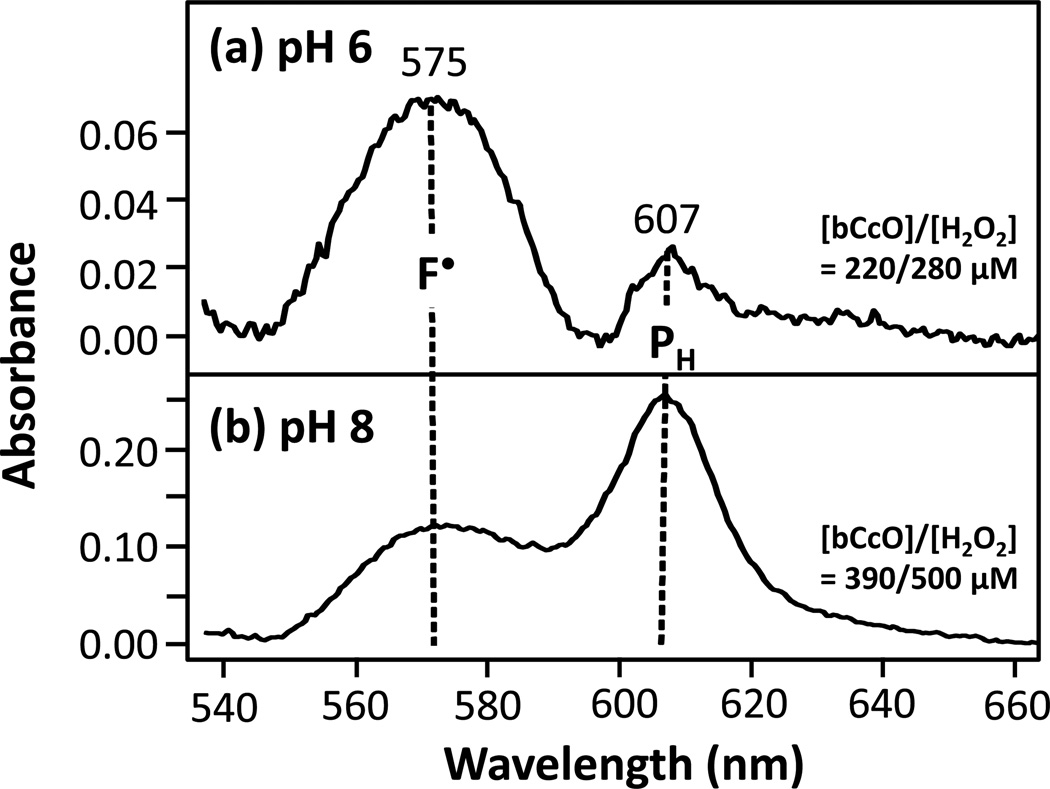
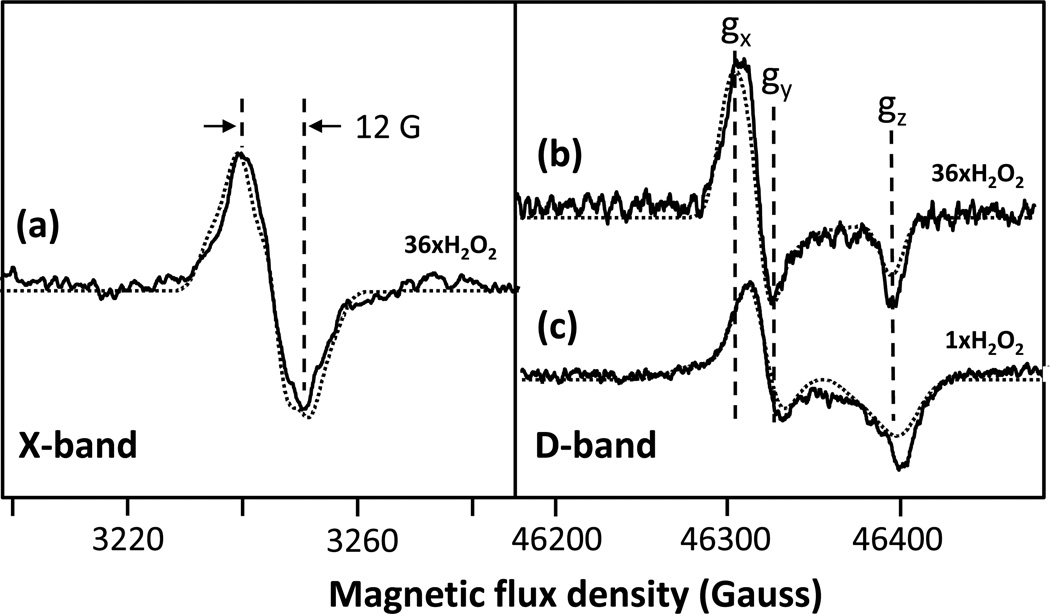
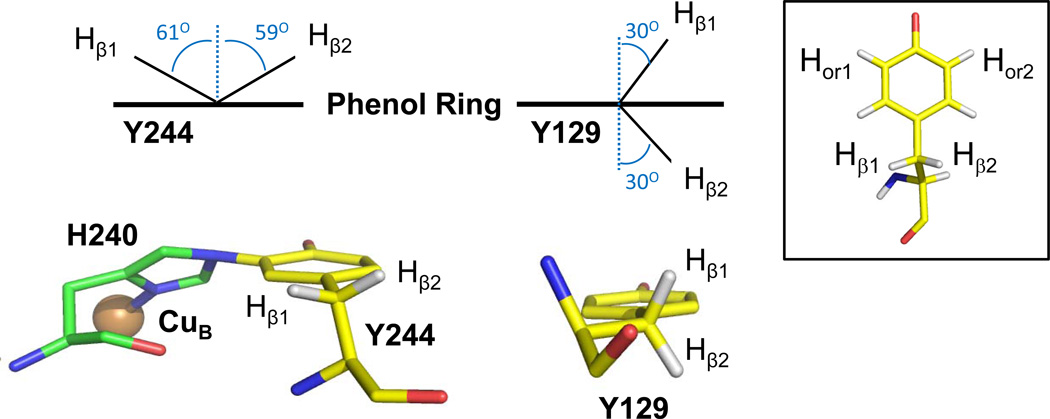
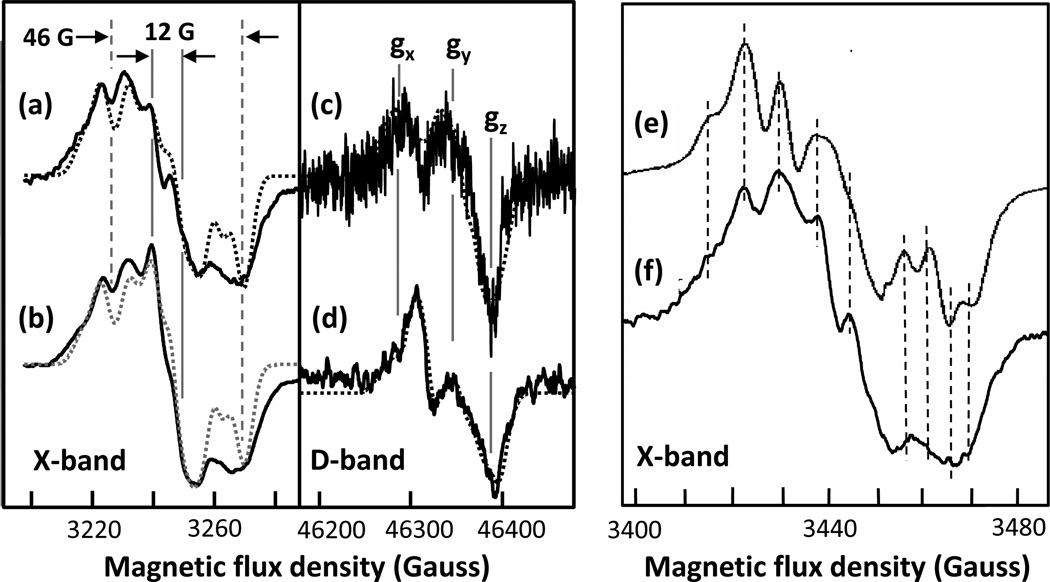
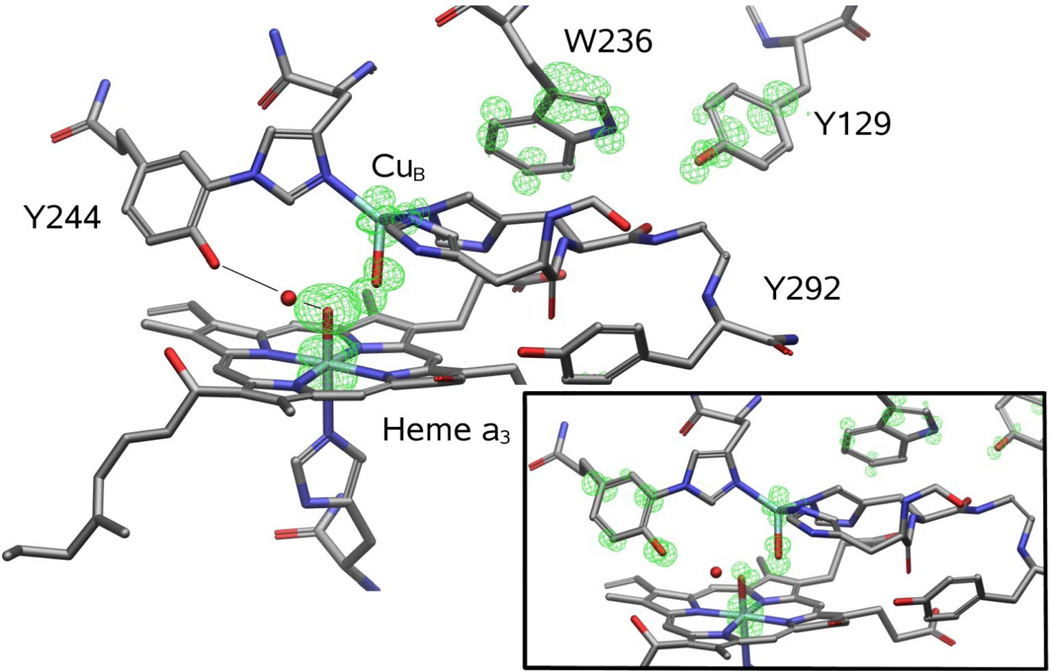
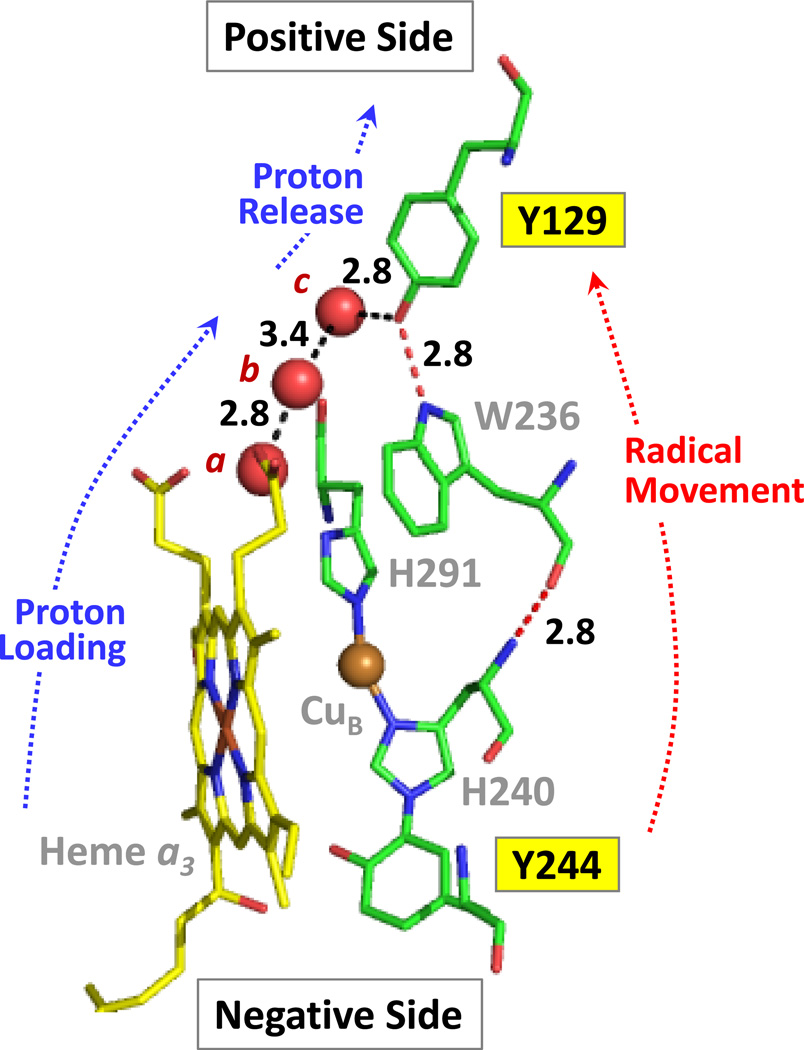
The reaction of oxidized bovine cytochrome c oxidase (bCcO) with hydrogen peroxide (H(2)O(2)) was studied by electron paramagnetic resonance (EPR) to determine the properties of radical intermediates. Two distinct radicals with widths of 12 and 46 G are directly observed by X-band EPR in the reaction of bCcO with H(2)O(2) at pH 6 and pH 8. High-frequency EPR (D-band) provides assignments to tyrosine for both radicals based on well-resolved g-tensors. The wide radical (46 G) exhibits g-values similar to a radical generated on L-Tyr by UV-irradiation and to tyrosyl radicals identified in many other enzyme systems. In contrast, the g-values of the narrow radical (12 G) deviate from L-Tyr in a trend akin to the radicals on tyrosines with substitutions at the ortho position. X-band EPR demonstrates that the two tyrosyl radicals differ in the orientation of their β-methylene protons. The 12 G wide radical has minimal hyperfine structure and can be fit using parameters unique to the post-translationally modified Y244 in bCcO. The 46 G wide radical has extensive hyperfine structure and can be fit with parameters consistent with Y129. The results are supported by mixed quantum mechanics and molecular mechanics calculations. In addition to providing spectroscopic evidence of a radical formed on the post-translationally modified tyrosine in CcO, this study resolves the much debated controversy of whether the wide radical seen at low pH in the bovine enzyme is a tyrosine or tryptophan. The possible role of radical formation and migration in proton translocation is discussed.
Figures
Similar articles
-
Radical formation in cytochrome c oxidase.Biochim Biophys Acta. 2011 Oct;1807(10):1295-304. doi: 10.1016/j.bbabio.2011.06.012. Epub 2011 Jun 22. Biochim Biophys Acta. 2011. PMID: 21718686 Free PMC article.
-
Tryptophan or tyrosine? On the nature of the amino acid radical formed following hydrogen peroxide treatment of cytochrome c oxidase.Biochim Biophys Acta. 2004 Apr 12;1655(1-3):372-80. doi: 10.1016/j.bbabio.2003.06.006. Biochim Biophys Acta. 2004. PMID: 15100053 Review.
-
Reaction of bovine cytochrome c oxidase with hydrogen peroxide produces a tryptophan cation radical and a porphyrin cation radical.Biochemistry. 2000 May 23;39(20):5921-8. doi: 10.1021/bi992614q. Biochemistry. 2000. PMID: 10821663
-
Direct evidence for a tyrosine radical in the reaction of cytochrome c oxidase with hydrogen peroxide.Biochemistry. 1999 Jul 20;38(29):9179-84. doi: 10.1021/bi9911987. Biochemistry. 1999. PMID: 10413492
-
Important roles of tyrosines in photosystem II and cytochrome oxidase.Biochim Biophys Acta. 2004 Apr 12;1655(1-3):45-50. doi: 10.1016/j.bbabio.2003.07.003. Biochim Biophys Acta. 2004. PMID: 15100015 Review.
Cited by
-
Proton translocation in cytochrome c oxidase: insights from proton exchange kinetics and vibrational spectroscopy.Biochim Biophys Acta. 2015 Jan;1847(1):98-108. doi: 10.1016/j.bbabio.2014.09.008. Epub 2014 Sep 28. Biochim Biophys Acta. 2015. PMID: 25268561 Free PMC article. Review.
-
Formal Reduction Potentials of Difluorotyrosine and Trifluorotyrosine Protein Residues: Defining the Thermodynamics of Multistep Radical Transfer.J Am Chem Soc. 2017 Mar 1;139(8):2994-3004. doi: 10.1021/jacs.6b11011. Epub 2017 Feb 21. J Am Chem Soc. 2017. PMID: 28171730 Free PMC article.
-
Tryptophan Can Promote Oxygen Reduction to Water in a Biosynthetic Model of Heme Copper Oxidases.Biochemistry. 2023 Jan 17;62(2):388-395. doi: 10.1021/acs.biochem.2c00300. Epub 2022 Oct 10. Biochemistry. 2023. PMID: 36215733 Free PMC article.
-
Coupled transport of electrons and protons in a bacterial cytochrome c oxidase-DFT calculated properties compared to structures and spectroscopies.Phys Chem Chem Phys. 2020 Dec 7;22(46):26652-26668. doi: 10.1039/d0cp04848h. Phys Chem Chem Phys. 2020. PMID: 33231596 Free PMC article.
-
Direct EPR observation of a tyrosyl radical in a functional oxidase model in myoglobin during both H2O2 and O2 reactions.J Am Chem Soc. 2014 Jan 29;136(4):1174-1177. doi: 10.1021/ja4091885. Epub 2014 Jan 14. J Am Chem Soc. 2014. PMID: 24383850 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
