

Noradrenergic modulation of wakefulness/arousal
- PMID: 22296742
- PMCID: PMC3278579
- DOI: 10.1016/j.smrv.2011.12.003
Noradrenergic modulation of wakefulness/arousal
Abstract
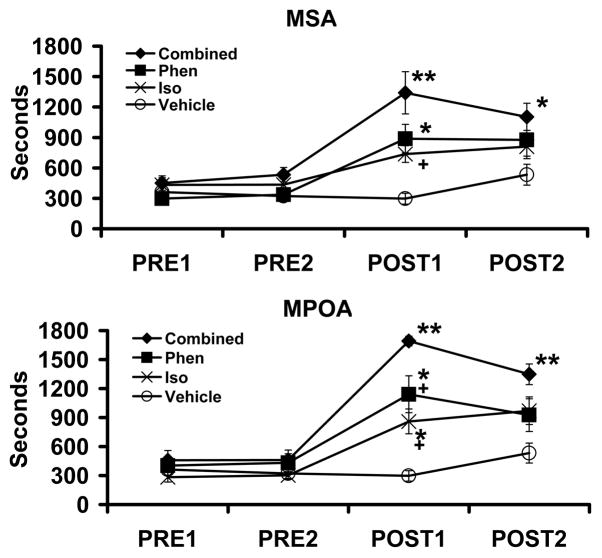
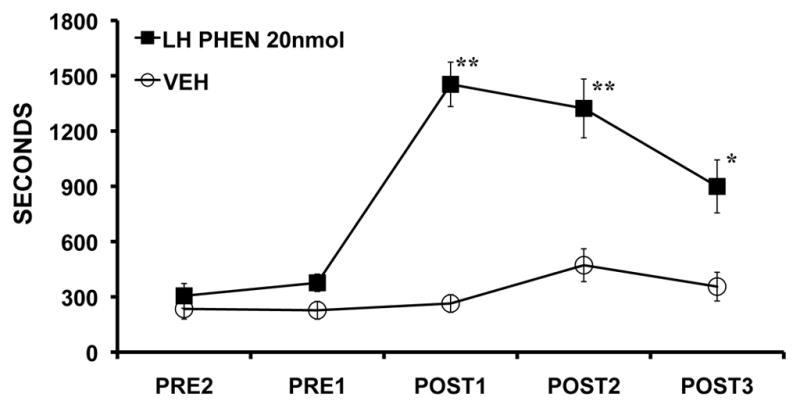
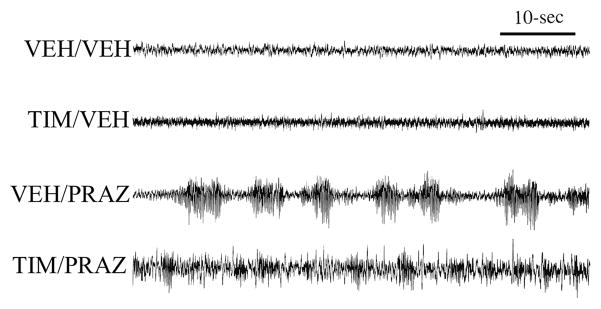
The locus coeruleus-noradrenergic system supplies norepinephrine throughout the central nervous system. State-dependent neuronal discharge activity of locus coeruleus noradrenergic neurons has long-suggested a role of this system in the induction of an alert waking state. Work over the past two decades provides unambiguous evidence that the locus coeruleus, and likely other noradrenergic nuclei, exert potent wake-promoting actions via an activation of noradrenergic β- and α₁-receptors located within multiple subcortical structures, including the general regions of the medial septal area, the medial preoptic area and, most recently, the lateral hypothalamus. Conversely, global blockade of β- and α₁-receptors or suppression of norepinephrine release results in profound sedation. The wake-promoting action of central noradrenergic neurotransmission has clinical implications for treatment of sleep/arousal disorders, such as insomnia and narcolepsy, and clinical conditions associated with excessive arousal, such as post-traumatic stress disorder.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
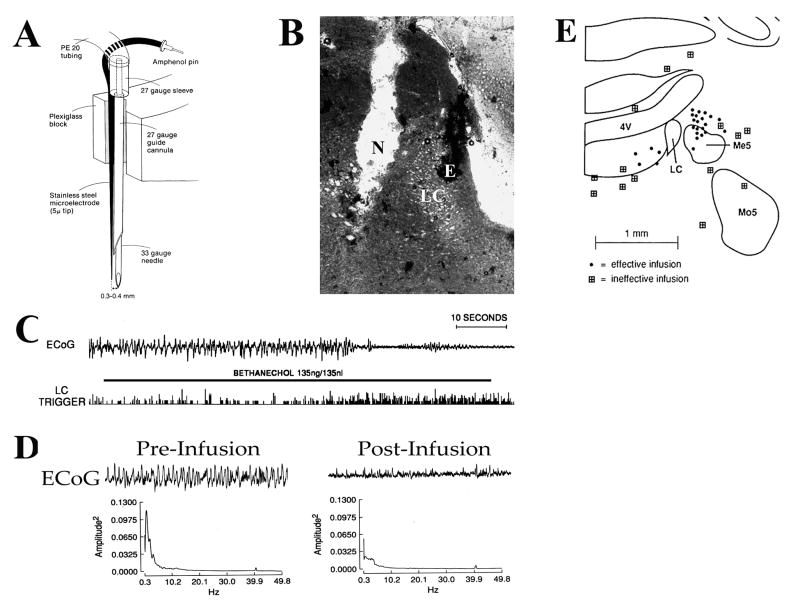
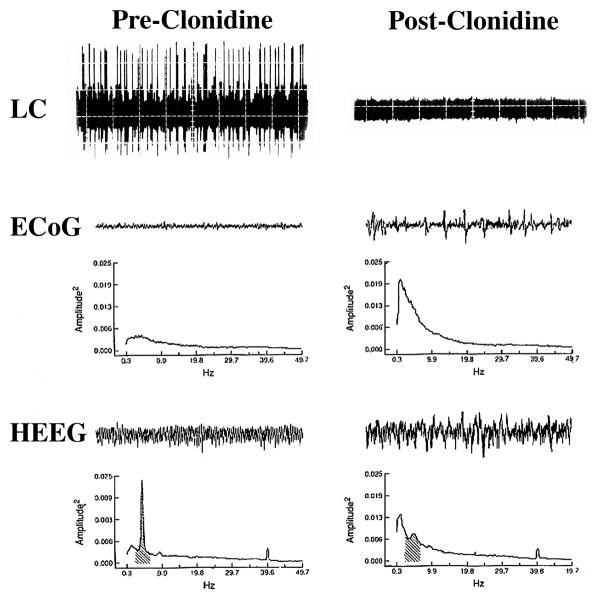
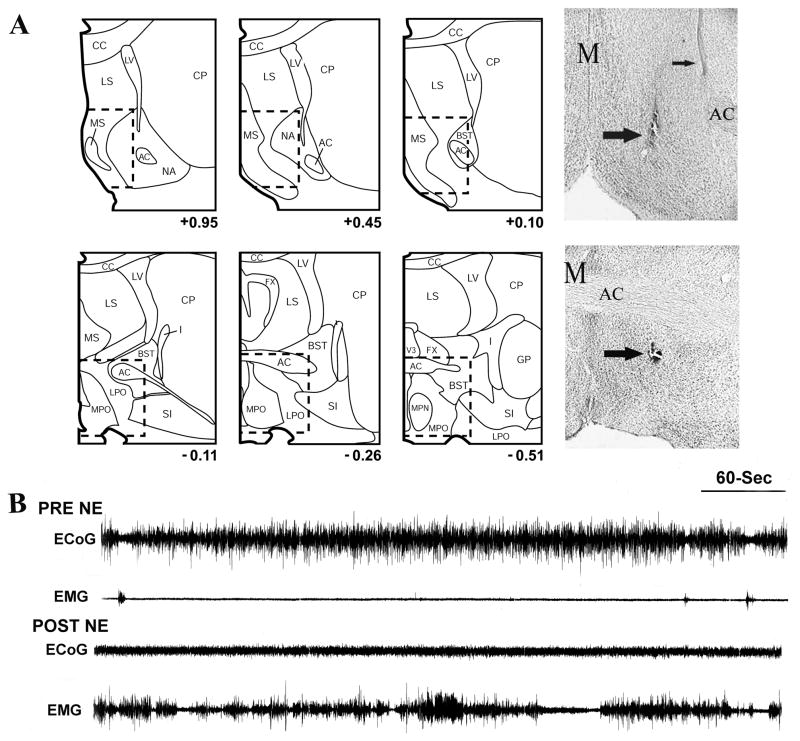
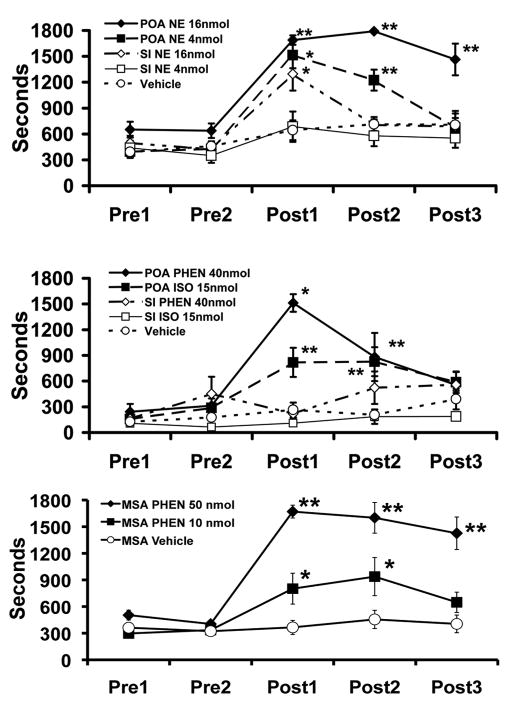
References
-
- Bushnell MC, Goldberg ME, Robinson DL. Behavioral enhancement of visual responses in monkey cerebral cortex. I. Modulation in posterior parietal cortex related to selective visual attention. J Neurophysiol. 1981;46(4):755–72. - PubMed
-
- Evarts EV. Effects of sleep and waking on spontaneous and evoked discharge of single units in visual cortex. Fed Proc. 1960;19:828–37. - PubMed
-
- Von Economo C. Sleep as a problem of localization. J Nerv Ment Dis. 1930;71:249–59.
-
- Bremer F. Cerebral activity during sleep and narcosis: Contribution to the study of the mechanisms of sleep. Bull l’Acad Royale Med Belgique. 1937;4:240–75.
-
- Moruzzi G, Magoun HW. Brain stem reticular formation and activation of the EEG. Electroencephalogr Clin Neurophysiol. 1949;1:455–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
