

Fast synaptic inhibition in spinal sensory processing and pain control
- PMID: 22298656
- PMCID: PMC3590010
- DOI: 10.1152/physrev.00043.2010
Fast synaptic inhibition in spinal sensory processing and pain control
Abstract
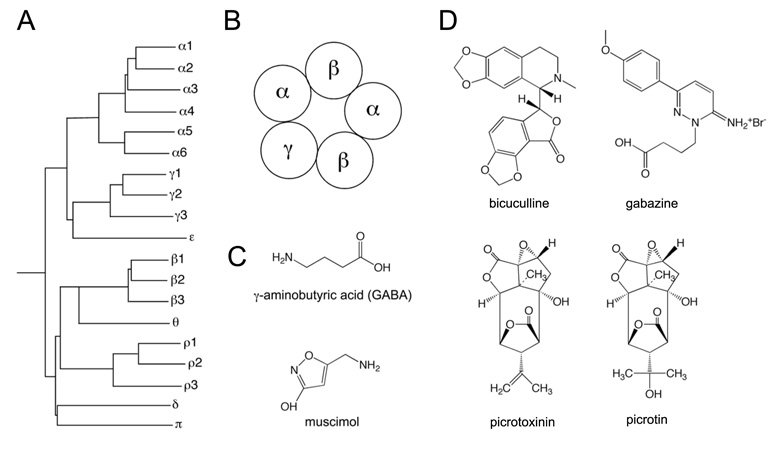
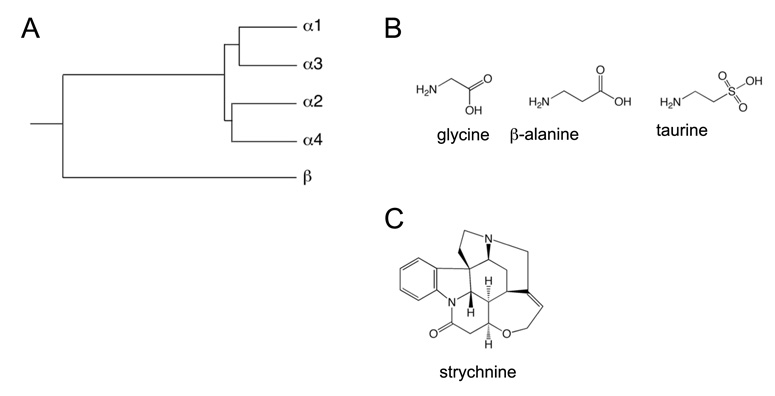
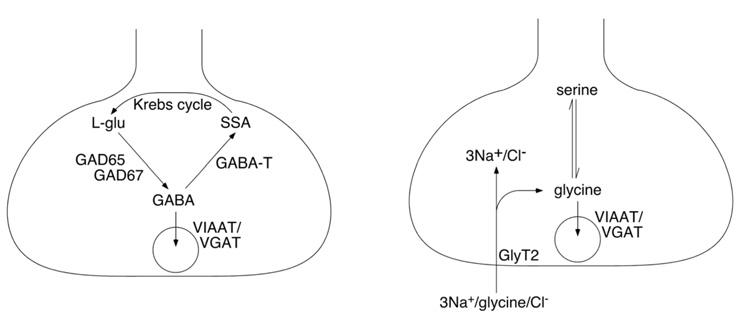
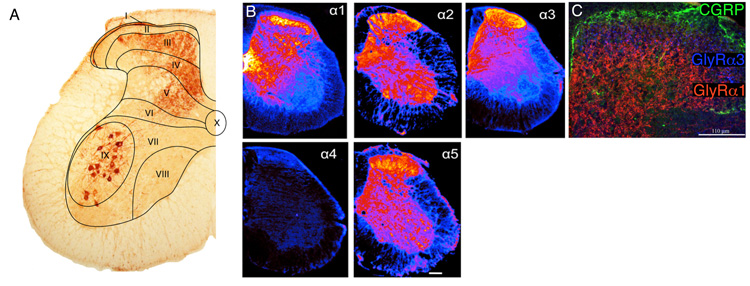
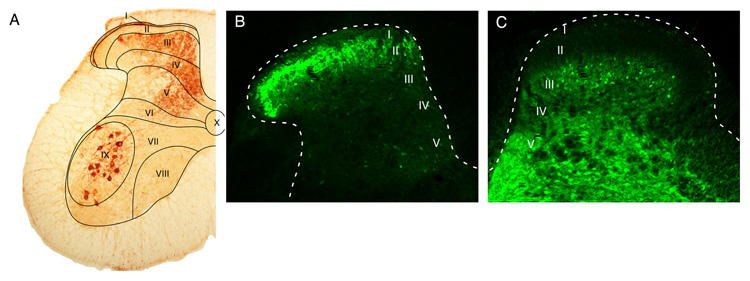
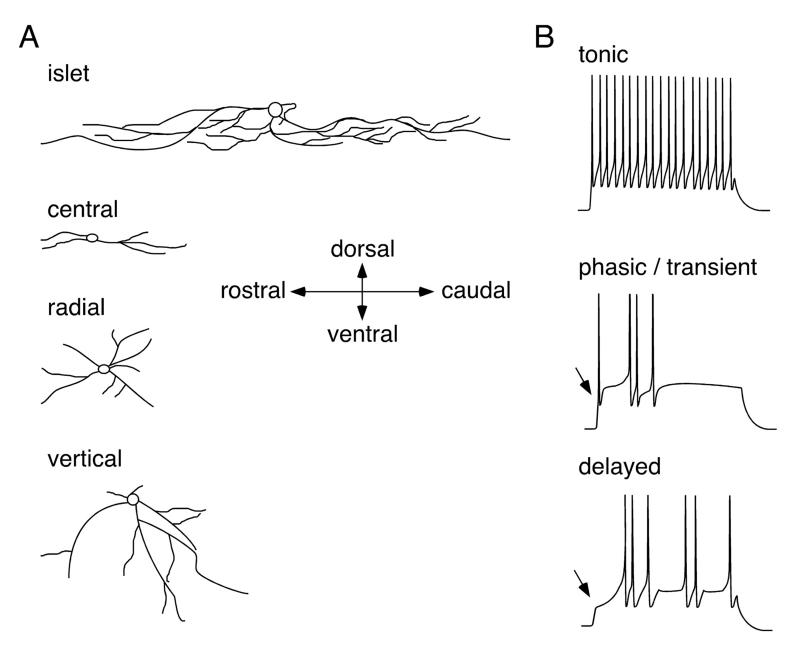
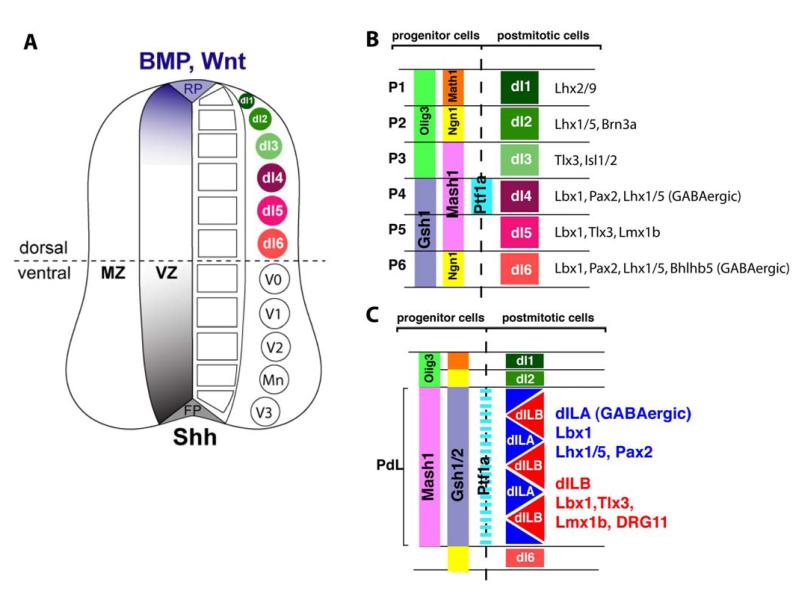
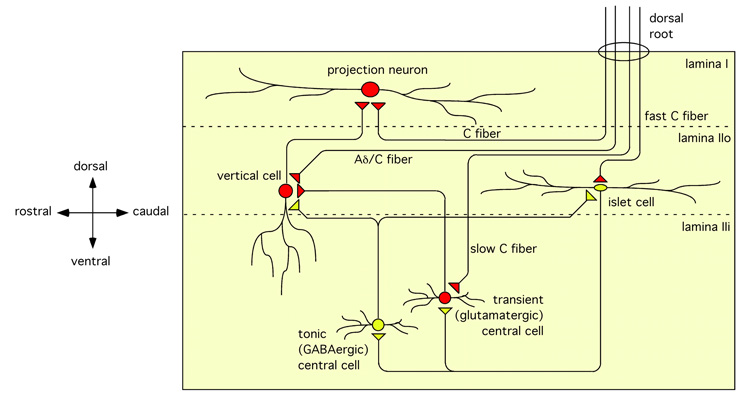
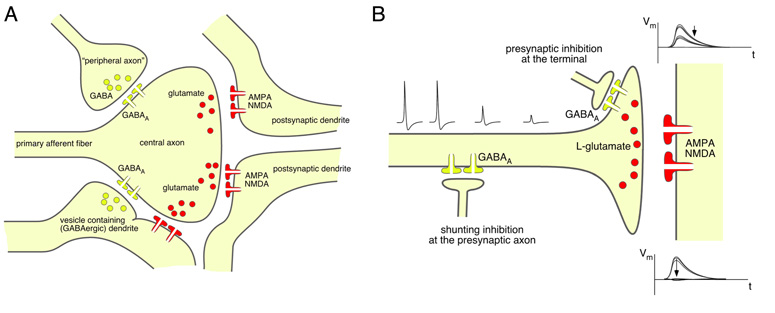
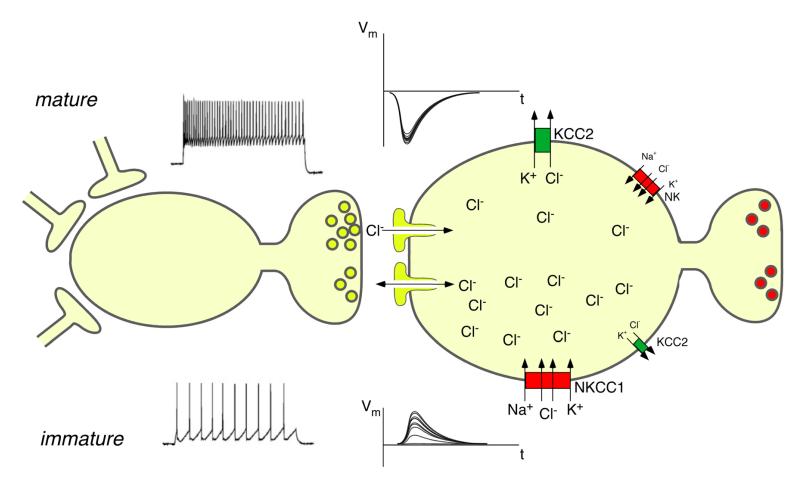
The two amino acids GABA and glycine mediate fast inhibitory neurotransmission in different CNS areas and serve pivotal roles in the spinal sensory processing. Under healthy conditions, they limit the excitability of spinal terminals of primary sensory nerve fibers and of intrinsic dorsal horn neurons through pre- and postsynaptic mechanisms, and thereby facilitate the spatial and temporal discrimination of sensory stimuli. Removal of fast inhibition not only reduces the fidelity of normal sensory processing but also provokes symptoms very much reminiscent of pathological and chronic pain syndromes. This review summarizes our knowledge of the molecular bases of spinal inhibitory neurotransmission and its organization in dorsal horn sensory circuits. Particular emphasis is placed on the role and mechanisms of spinal inhibitory malfunction in inflammatory and neuropathic chronic pain syndromes.
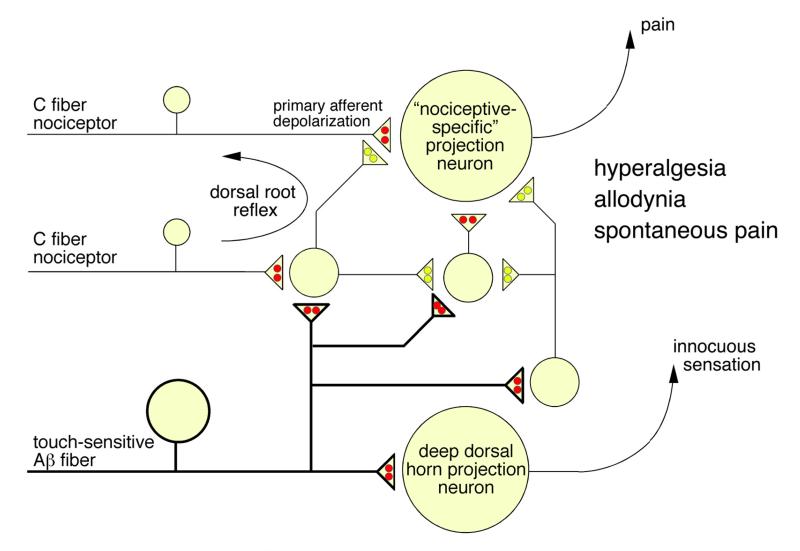
Figures
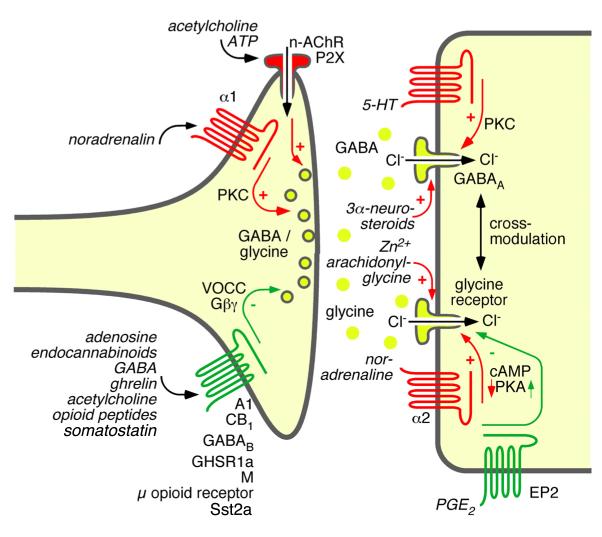
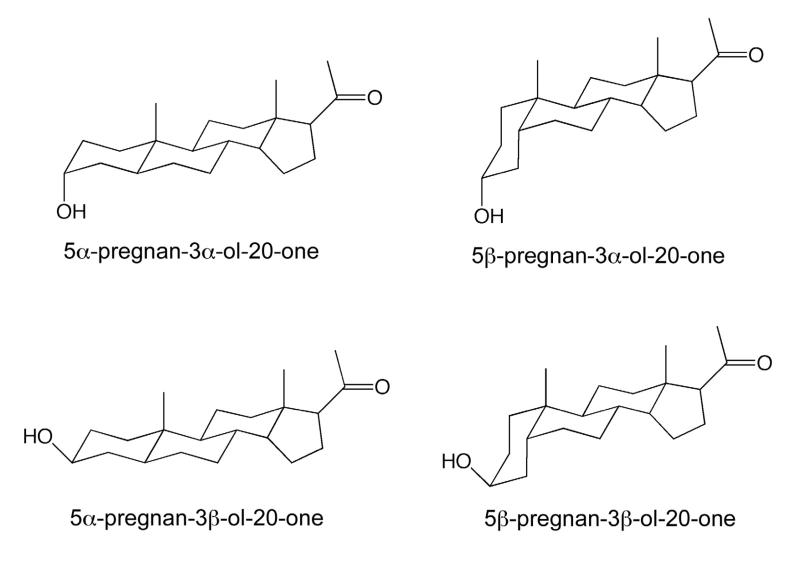
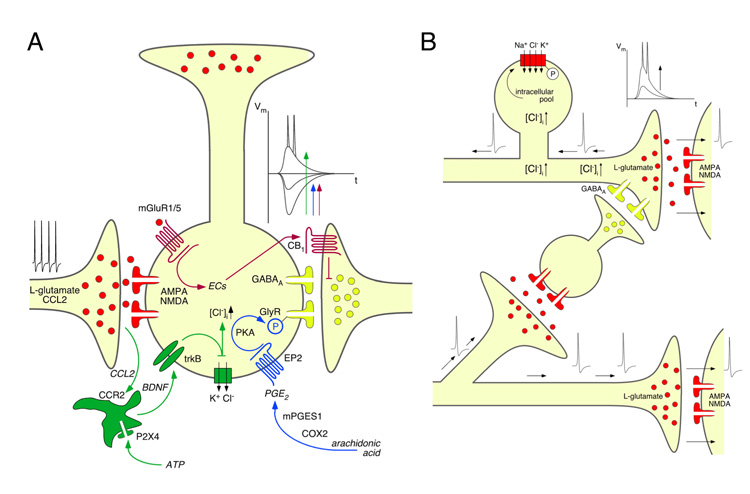
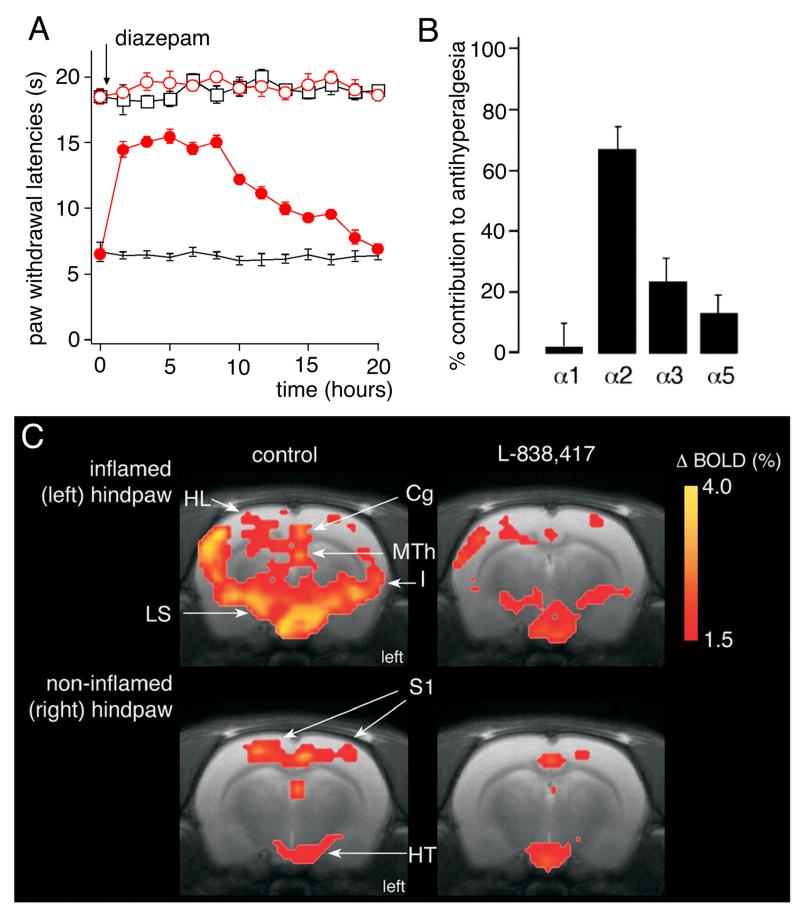
References
-
- Abe K, Kato G, Katafuchi T, Tamae A, Furue H, Yoshimura M. Responses to 5-HT in morphologically identified neurons in the rat substantia gelatinosa in vitro. Neuroscience. 2009;159:316–324. - PubMed
-
- Abrahamsen B, Zhao J, Asante CO, Cendan CM, Marsh S, Martinez-Barbera JP, Nassar MA, Dickenson AH, Wood JN. The cell and molecular basis of mechanical, cold, and inflammatory pain. Science. 2008;321:702–705. - PubMed
-
- Aguayo LG, Peoples RW, Yeh HH, Yevenes GE. GABAA receptors as molecular sites of ethanol action. Direct or indirect actions? Curr Top Med Chem. 2002;2:869–885. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical