

BMP-9 induced endothelial cell tubule formation and inhibition of migration involves Smad1 driven endothelin-1 production
- PMID: 22299030
- PMCID: PMC3267722
- DOI: 10.1371/journal.pone.0030075
BMP-9 induced endothelial cell tubule formation and inhibition of migration involves Smad1 driven endothelin-1 production
Abstract
Background: Bone morphogenetic proteins (BMPs) and their receptors, such as bone morphogenetic protein receptor (BMPR) II, have been implicated in a wide variety of disorders including pulmonary arterial hypertension (PAH). Similarly, endothelin-1 (ET-1), a mitogen and vasoconstrictor, is upregulated in PAH and endothelin receptor antagonists are used in its treatment. We sought to determine whether there is crosstalk between BMP signalling and the ET-1 axis in human pulmonary artery endothelial cells (HPAECs), possible mechanisms involved in such crosstalk and functional consequences thereof.
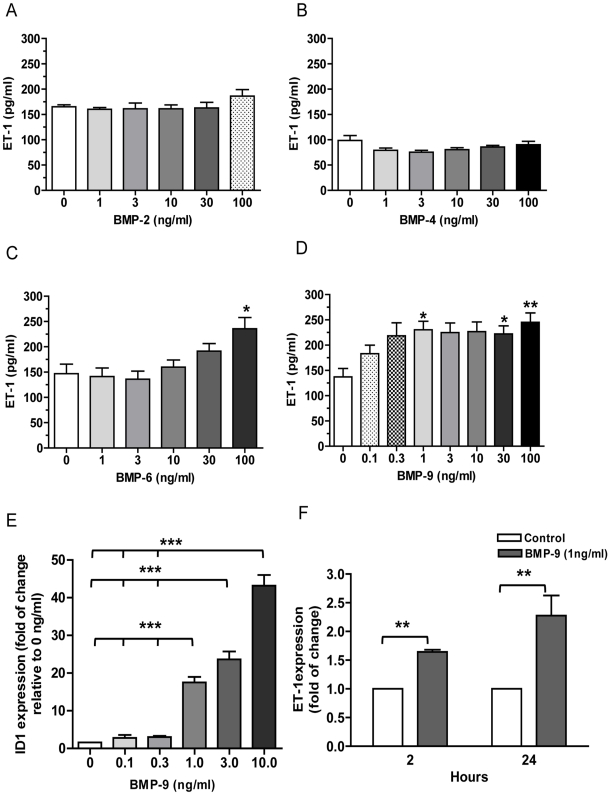
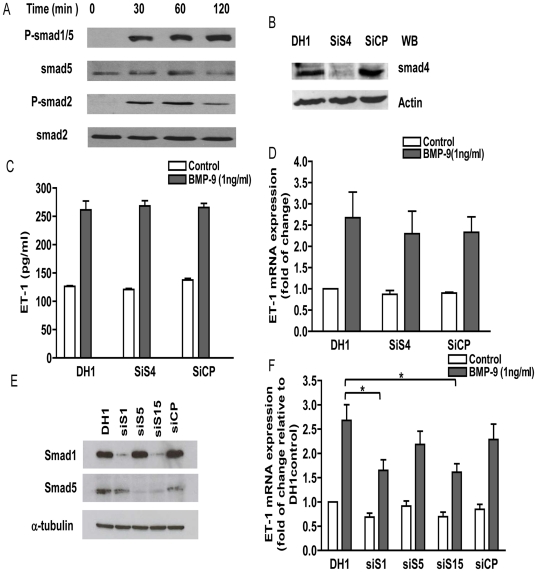
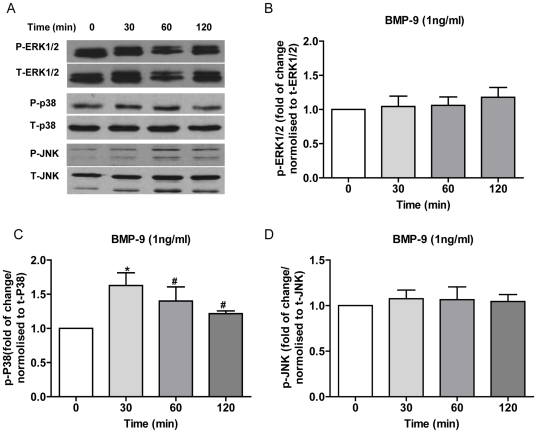
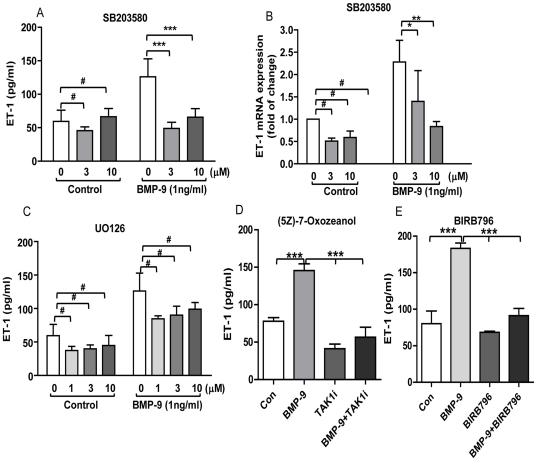
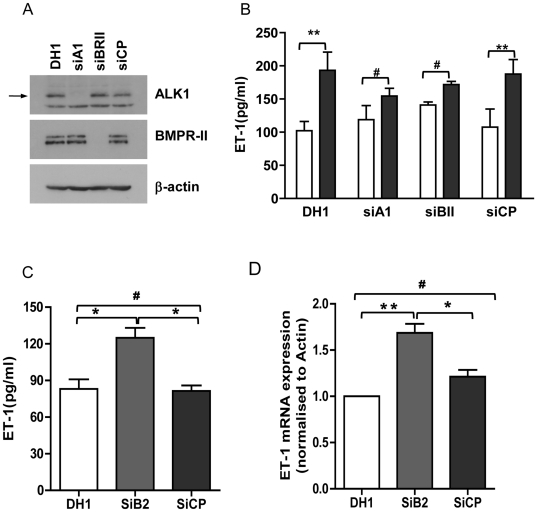
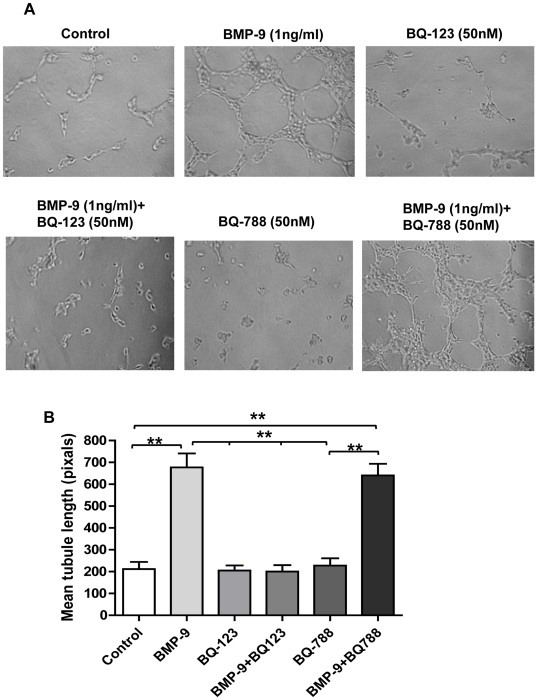
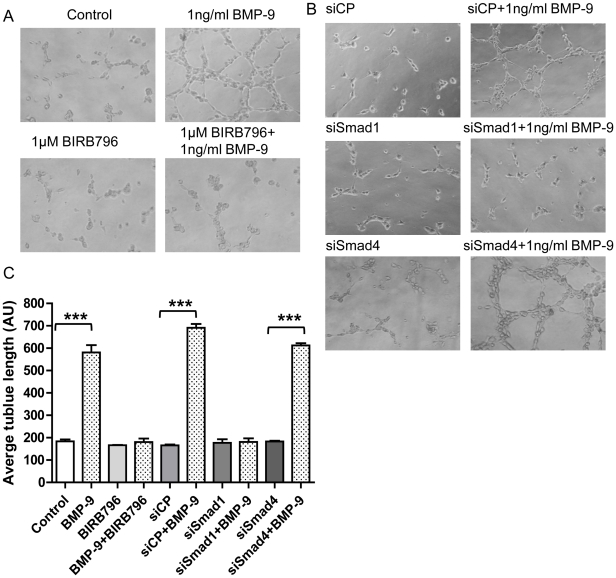
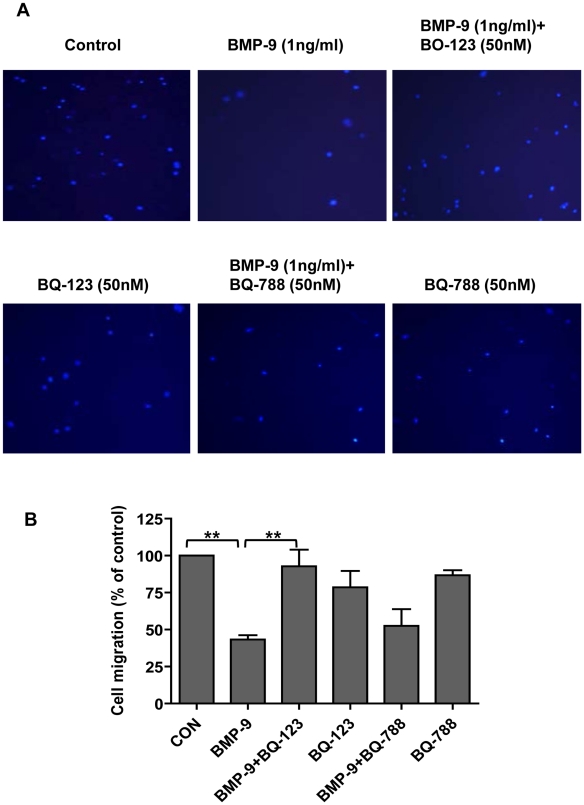
Methodology/principal finding: Using western blot, real time RT-PCR, ELISA and small RNA interference methods we provide evidence that in HPAECs BMP-9, but not BMP-2, -4 and -6 significantly stimulated ET-1 release under physiological concentrations. This release is mediated by both Smad1 and p38 MAPK and is independent of the canonical Smad4 pathway. Moreover, knocking down the ALK1 receptor or BMPR II attenuates BMP-9 stimulated ET-1 release, whilst causing a significant increase in prepro ET-1 mRNA transcription and mature peptide release. Finally, BMP-9 induced ET-1 release is involved in both inhibition of endothelial cell migration and promotion of tubule formation.
Conclusions/significance: Although our data does not support an important role for BMP-9 as a source of increased endothelial ET-1 production seen in human PAH, BMP-9 stimulated ET-1 production is likely to be important in angiogenesis and vascular stability. However, increased ET-1 production by endothelial cells as a consequence of BMPR II dysfunction may be clinically relevant in the pathogenesis of PAH.
Conflict of interest statement
Figures
References
-
- Zeng S, Chen J, Shen H. Controlling of bone morphogenetic protein signaling. Cell Signal. 2010;22:888–893. - PubMed
-
- Kimura N, Matsuo R, Shibuya H, Nakashima K, Taga T. BMP2-induced apoptosis is mediated by activation of the TAK1-p38 kinase pathway that is negatively regulated by Smad6. J Biol Chem. 2000;275:17647–17652. - PubMed
-
- Su D, Peng X, Zhu S, Huang Y, Dong Z, et al. Role of p38 MAPK pathway in BMP4-mediated Smad-dependent premature senescence in lung cancer cells. Biochem J. 2010;433:333–343. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
