

Disulfide linkages mediating nucleocapsid protein dimerization are not required for porcine arterivirus infectivity
- PMID: 22301142
- PMCID: PMC3318635
- DOI: 10.1128/JVI.06709-11
Disulfide linkages mediating nucleocapsid protein dimerization are not required for porcine arterivirus infectivity
Abstract
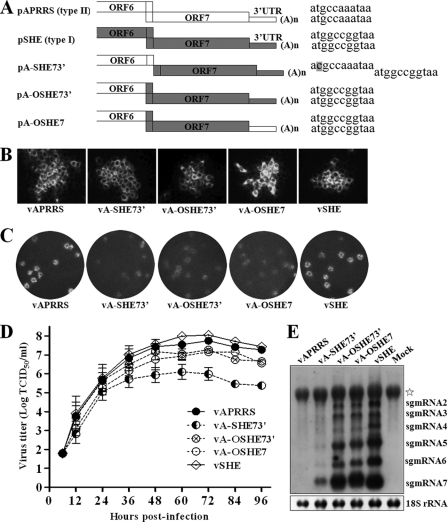
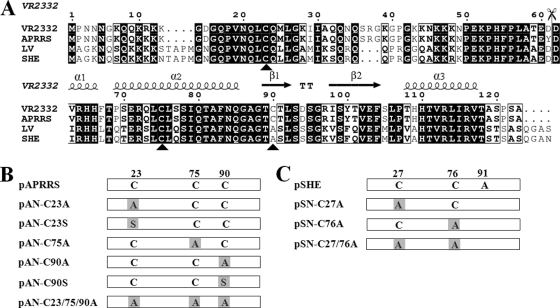
The nucleocapsid (N) proteins of the North American (type II) and European (type I) genotypes of porcine reproductive and respiratory syndrome virus (PRRSV) share only approximately 60% genetic identity, and the functionality of N in both genotypes, especially its role in virion assembly, is still poorly understood. In this study, we demonstrated that the ORF7 3' untranslated region or ORF7 of type I is functional in the type II PRRSV background. Based on these results, we postulated that the cysteine at position 90 (Cys90) of the type II N protein, which corresponds to an alanine in the type I protein, is nonessential for virus infectivity. The replacement of Cys90 with alanine confirmed this hypothesis. We then hypothesized that all of the cysteines in the N protein could be replaced by alanines. Mutational analysis revealed that, in contradiction to previously reported findings, the replacement of all of the cysteines, either singly or in combination, did not impair the growth of either type II or type I PRRSV. Treatment with the alkylating agent N-ethylmaleimide inhibited cysteine-mediated N dimerization in living cells but not in released virions. Additionally, bimolecular fluorescence complementation assays revealed noncovalent interactions in living cells among the N and C termini and between the N-terminal and C-terminal regions of the N proteins of both genotypes of PRRSV. These results demonstrate that the disulfide linkages mediating the N dimerization are not required for PRRSV viability and help to promote our understanding of the mechanism underlying arterivirus particle assembly.
Figures
Similar articles
-
Homo-oligomerization of the porcine reproductive and respiratory syndrome virus nucleocapsid protein and the role of disulfide linkages.J Virol. 2003 Apr;77(8):4546-57. doi: 10.1128/jvi.77.8.4546-4557.2003. J Virol. 2003. PMID: 12663761 Free PMC article.
-
A DNA-launched reverse genetics system for porcine reproductive and respiratory syndrome virus reveals that homodimerization of the nucleocapsid protein is essential for virus infectivity.Virology. 2005 Jan 5;331(1):47-62. doi: 10.1016/j.virol.2004.10.026. Virology. 2005. PMID: 15582652
-
An SH3 binding motif within the nucleocapsid protein of porcine reproductive and respiratory syndrome virus interacts with the host cellular signaling proteins STAMI, TXK, Fyn, Hck, and cortactin.Virus Res. 2015 Jun 2;204:31-9. doi: 10.1016/j.virusres.2015.04.004. Epub 2015 Apr 13. Virus Res. 2015. PMID: 25882913
-
Current knowledge on the structural proteins of porcine reproductive and respiratory syndrome (PRRS) virus: comparison of the North American and European isolates.Arch Virol. 2000;145(4):659-88. doi: 10.1007/s007050050662. Arch Virol. 2000. PMID: 10893147 Free PMC article. Review.
-
Identification of dispensable nucleotide sequence in 3' untranslated region of porcine reproductive and respiratory syndrome virus.Virus Res. 2010 Dec;154(1-2):38-47. doi: 10.1016/j.virusres.2010.08.027. Epub 2010 Sep 15. Virus Res. 2010. PMID: 20833212 Free PMC article. Review.
Cited by
-
Porcine Reproductive and Respiratory Syndrome Virus Reverse Genetics and the Major Applications.Viruses. 2020 Oct 31;12(11):1245. doi: 10.3390/v12111245. Viruses. 2020. PMID: 33142752 Free PMC article. Review.
-
The Nsp12-coding region of type 2 PRRSV is required for viral subgenomic mRNA synthesis.Emerg Microbes Infect. 2019;8(1):1501-1510. doi: 10.1080/22221751.2019.1679010. Emerg Microbes Infect. 2019. PMID: 31631782 Free PMC article.
-
A novel porcine reproductive and respiratory syndrome virus vector system that stably expresses enhanced green fluorescent protein as a separate transcription unit.Vet Res. 2013 Oct 31;44(1):104. doi: 10.1186/1297-9716-44-104. Vet Res. 2013. PMID: 24176053 Free PMC article.
-
DExD/H-Box Helicase 36 Signaling via Myeloid Differentiation Primary Response Gene 88 Contributes to NF-κB Activation to Type 2 Porcine Reproductive and Respiratory Syndrome Virus Infection.Front Immunol. 2017 Oct 23;8:1365. doi: 10.3389/fimmu.2017.01365. eCollection 2017. Front Immunol. 2017. PMID: 29123520 Free PMC article.
-
Nuclear localization signal in TRIM22 is essential for inhibition of type 2 porcine reproductive and respiratory syndrome virus replication in MARC-145 cells.Virus Genes. 2019 Oct;55(5):660-672. doi: 10.1007/s11262-019-01691-x. Epub 2019 Aug 2. Virus Genes. 2019. PMID: 31375995 Free PMC article.
References
-
- Allende R, et al. 1999. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J. Gen. Virol. 80:307–315 - PubMed
-
- Aparicio F, Sanchez-Navarro JA, Pallas V. 2006. In vitro and in vivo mapping of the Prunus necrotic ringspot virus coat protein C-terminal dimerization domain by bimolecular fluorescence complementation. J. Gen. Virol. 87:1745–1750 - PubMed
-
- Benfield DA, et al. 1992. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 4:127–133 - PubMed
-
- Boyko V, et al. 2006. Coassembly and complementation of Gag proteins from HIV-1 and HIV-2, two distinct human pathogens. Mol. Cell 23:281–287 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
