

Continual low-level MEK inhibition ameliorates cardio-facio-cutaneous phenotypes in zebrafish
- PMID: 22301711
- PMCID: PMC3380717
- DOI: 10.1242/dmm.008672
Continual low-level MEK inhibition ameliorates cardio-facio-cutaneous phenotypes in zebrafish
Abstract
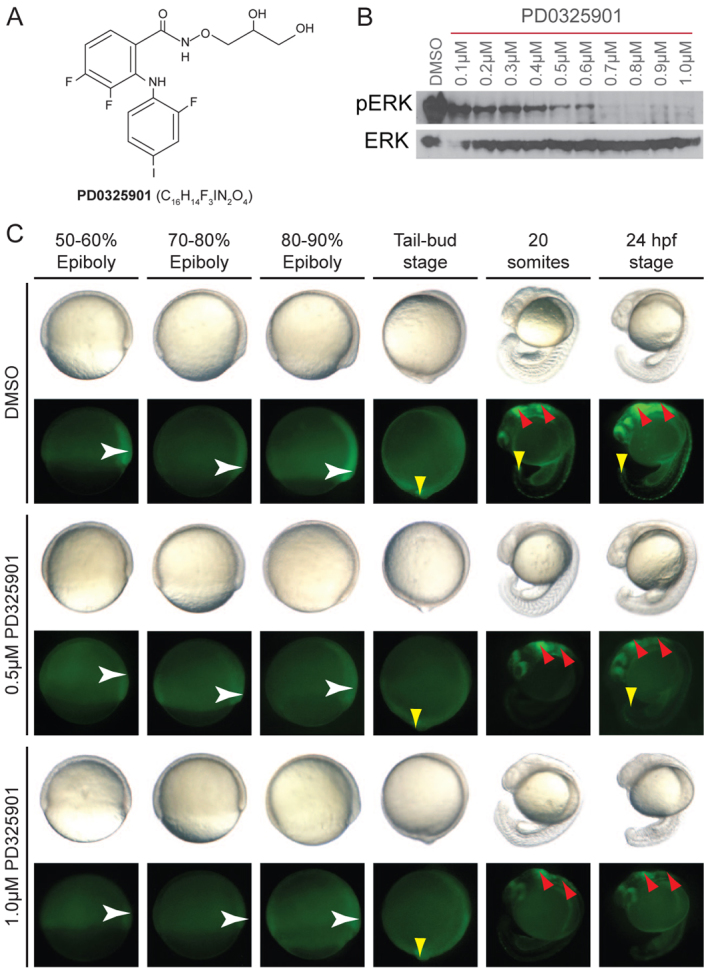
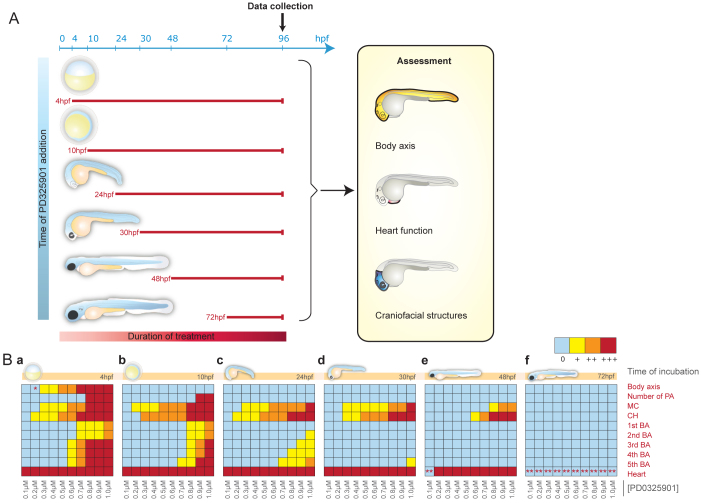
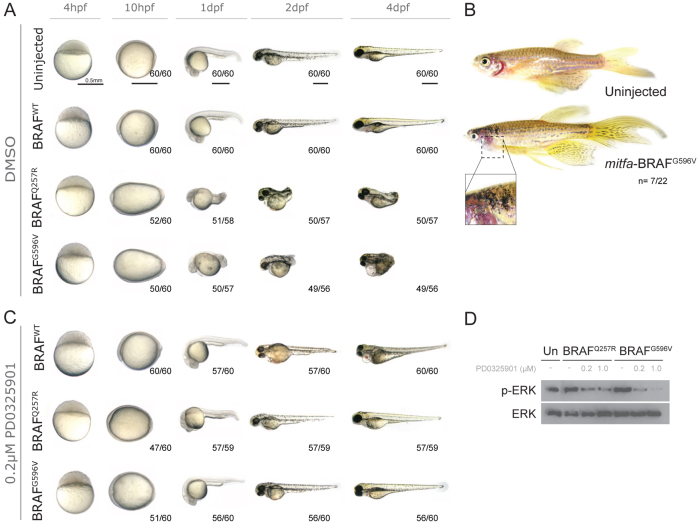
Cardio-facio-cutaneous (CFC) syndrome is caused by germline mutations in KRAS, BRAF and MEK1/2. The highly selective and potent MEK inhibitors that have been developed as anti-cancer agents hold potential as therapeutics for CFC syndrome. We have previously shown that the effects of CFC mutations on zebrafish gastrulation can be prevented by a 1-hour treatment with MEK inhibitors within a specific developmental time-window. However, MEK activity is essential for normal development and PD0325901 treatment outside this treatment window leads to additional developmental defects in MEK-dependent tissues. We now test ten different doses of PD0325901 at six developmental time points and assess the effects on body axis length, heart development and craniofacial structures in zebrafish embryos. Notably, we find that a continuous low-level dose of PD0325901 that has only minor inhibition of MEK activity can prevent the action of both the common CFC BRAF(Q257R) kinase-active allele and the BRAF(G596V) kinase-impaired mutant allele through the first 5 days of development. These results provide a detailed study of the effects of PD0325901 in development and show that, unlike in cancer, which requires robust inhibition of MAPK signalling, a partial reduction in phospho-ERK1/2 activity is sufficient to moderate the developmental effects of BRAF(CFC) mutations.
Figures
Similar articles
-
New BRAF knockin mice provide a pathogenetic mechanism of developmental defects and a therapeutic approach in cardio-facio-cutaneous syndrome.Hum Mol Genet. 2014 Dec 15;23(24):6553-66. doi: 10.1093/hmg/ddu376. Epub 2014 Jul 17. Hum Mol Genet. 2014. PMID: 25035421
-
Activated Braf induces esophageal dilation and gastric epithelial hyperplasia in mice.Hum Mol Genet. 2017 Dec 1;26(23):4715-4727. doi: 10.1093/hmg/ddx354. Hum Mol Genet. 2017. PMID: 28973166
-
Constitutive activation of B-Raf in the mouse germ line provides a model for human cardio-facio-cutaneous syndrome.Proc Natl Acad Sci U S A. 2011 Mar 22;108(12):5015-20. doi: 10.1073/pnas.1016933108. Epub 2011 Mar 7. Proc Natl Acad Sci U S A. 2011. PMID: 21383153 Free PMC article.
-
Fibrous dysplasia in cardio-facio-cutaneous syndrome: A case report and review of literature.Am J Med Genet A. 2022 Sep;188(9):2732-2737. doi: 10.1002/ajmg.a.62879. Epub 2022 Jul 7. Am J Med Genet A. 2022. PMID: 35801299 Review.
-
LYMPHODYSPLASIA AND KRAS MUTATION: A CASE REPORT AND LITERATURE REVIEW.Lymphology. 2015 Sep;48(3):121-7. Lymphology. 2015. PMID: 26939159 Review.
Cited by
-
Abrogation of MAP4K4 protein function causes congenital anomalies in humans and zebrafish.Sci Adv. 2023 Apr 28;9(17):eade0631. doi: 10.1126/sciadv.ade0631. Epub 2023 Apr 26. Sci Adv. 2023. PMID: 37126546 Free PMC article.
-
Nodal signaling establishes a competency window for stochastic cell fate switching.Dev Cell. 2022 Dec 5;57(23):2604-2622.e5. doi: 10.1016/j.devcel.2022.11.008. Dev Cell. 2022. PMID: 36473458 Free PMC article.
-
RASopathies: unraveling mechanisms with animal models.Dis Model Mech. 2015 Aug 1;8(8):769-82. doi: 10.1242/dmm.020339. Dis Model Mech. 2015. PMID: 26203125 Free PMC article. Review.
-
Mutations in RIT1 cause Noonan syndrome - additional functional evidence and expanding the clinical phenotype.Clin Genet. 2016 Mar;89(3):359-66. doi: 10.1111/cge.12608. Epub 2015 Jun 4. Clin Genet. 2016. PMID: 25959749 Free PMC article.
-
How activating mutations affect MEK1 regulation and function.J Biol Chem. 2017 Nov 17;292(46):18814-18820. doi: 10.1074/jbc.C117.806067. Epub 2017 Oct 10. J Biol Chem. 2017. PMID: 29018093 Free PMC article.
References
-
- Chen P. C., Wakimoto H., Conner D., Araki T., Yuan T., Roberts A., Seidman C. E., Bronson R., Neel B. G., Seidman J. G., et al. (2010). Activation of multiple signaling pathways causes developmental defects in mice with a Noonan syndrome-associated Sos1 mutation. J. Clin. Invest. 120, 4353–4365 - PMC - PubMed
-
- Crump J. G., Maves L., Lawson N. D., Weinstein B. M., Kimmel C. B. (2004). An essential role for Fgfs in endodermal pouch formation influences later craniofacial skeletal patterning. Development 131, 5703–5716 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
