

Mice deficient in dual oxidase maturation factors are severely hypothyroid
- PMID: 22301785
- PMCID: PMC3286189
- DOI: 10.1210/me.2011-1320
Mice deficient in dual oxidase maturation factors are severely hypothyroid
Abstract
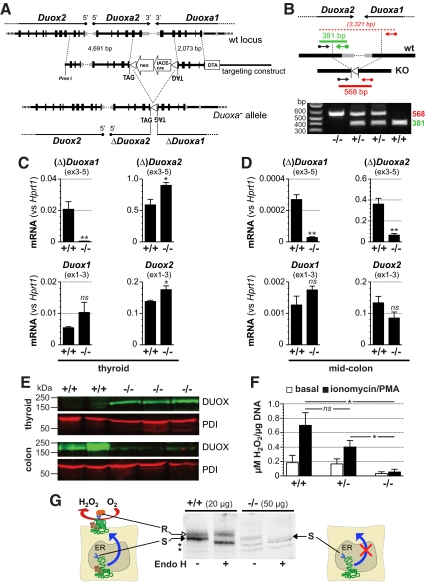
Dual oxidases (DUOX1 and DUOX2) are evolutionary conserved reduced nicotinamide adenine dinucleotide phosphate oxidases responsible for regulated hydrogen peroxide (H(2)O(2)) release of epithelial cells. Specific maturation factors (DUOXA1 and DUOXA2) are required for targeting of functional DUOX enzymes to the cell surface. Mutations in the single-copy Duox and Duoxa genes of invertebrates cause developmental defects with reduced survival, whereas knockdown in later life impairs intestinal epithelial immune homeostasis. In humans, mutations in both DUOX2 and DUOXA2 can cause congenital hypothyroidism with partial iodide organification defects compatible with a role of DUOX2-generated H(2)O(2) in driving thyroid peroxidase activity. The DUOX1/DUOXA1 system may account for residual iodide organification in patients with loss of DUOX2, but its physiological function is less clear. To provide a murine model recapitulating complete DUOX deficiency, we simultaneously targeted both Duoxa genes by homologous recombination. Knockout of Duoxa genes (Duoxa(-/-) mice) led to a maturation defect of DUOX proteins lacking Golgi processing of N-glycans and to loss of H(2)O(2) release from thyroid tissue. Postnatally, Duoxa(-/-) mice developed severe goitreous congenital hypothyroidism with undetectable serum T4 and maximally disinhibited TSH levels. Heterozygous mice had normal thyroid function parameters. (125)I uptake and discharge studies and probing of iodinated TG epitopes corroborated the iodide organification defect in Duoxa(-/-) mice. Duoxa(-/-) mice on continuous T4 replacement from P6 showed normal growth without an overt phenotype. Our results confirm in vivo the requirement of DUOXA for functional expression of DUOX-based reduced nicotinamide adenine dinucleotide phosphate oxidases and the role of DUOX isoenzymes as sole source of hormonogenic H(2)O(2).
Figures



Similar articles
-
Dual oxidases control release of hydrogen peroxide by the gastric epithelium to prevent Helicobacter felis infection and inflammation in mice.Gastroenterology. 2013 Nov;145(5):1045-54. doi: 10.1053/j.gastro.2013.07.011. Epub 2013 Jul 13. Gastroenterology. 2013. PMID: 23860501 Free PMC article.
-
A single copy of the recently identified dual oxidase maturation factor (DUOXA) 1 gene produces only mild transient hypothyroidism in a patient with a novel biallelic DUOXA2 mutation and monoallelic DUOXA1 deletion.J Clin Endocrinol Metab. 2011 May;96(5):E841-5. doi: 10.1210/jc.2010-2321. Epub 2011 Mar 2. J Clin Endocrinol Metab. 2011. PMID: 21367925 Free PMC article.
-
The Dual Oxidase Duox2 stabilized with DuoxA2 in an enzymatic complex at the surface of the cell produces extracellular H2O2 able to induce DNA damage in an inducible cellular model.Exp Cell Res. 2019 Nov 1;384(1):111620. doi: 10.1016/j.yexcr.2019.111620. Epub 2019 Sep 9. Exp Cell Res. 2019. PMID: 31513783
-
DUOX Defects and Their Roles in Congenital Hypothyroidism.Methods Mol Biol. 2019;1982:667-693. doi: 10.1007/978-1-4939-9424-3_37. Methods Mol Biol. 2019. PMID: 31172499 Review.
-
Genetic defects of hydrogen peroxide generation in the thyroid gland.J Endocrinol Invest. 2013 Apr;36(4):261-6. doi: 10.3275/8847. Epub 2013 Feb 12. J Endocrinol Invest. 2013. PMID: 23404134 Review.
Cited by
-
Pathogenesis of Multinodular Goiter in Elderly XB130-Deficient Mice: Alteration of Thyroperoxidase Affinity with Iodide and Hydrogen Peroxide.Thyroid. 2022 Apr;32(4):385-396. doi: 10.1089/thy.2021.0458. Thyroid. 2022. PMID: 34915750 Free PMC article.
-
Pluripotent stem cell differentiation reveals distinct developmental pathways regulating lung- versus thyroid-lineage specification.Development. 2017 Nov 1;144(21):3879-3893. doi: 10.1242/dev.150193. Epub 2017 Sep 25. Development. 2017. PMID: 28947536 Free PMC article.
-
Genetic variation in thyroid folliculogenesis influences susceptibility to hypothyroidism-induced hearing impairment.Mamm Genome. 2019 Feb;30(1-2):5-22. doi: 10.1007/s00335-019-09792-6. Epub 2019 Feb 18. Mamm Genome. 2019. PMID: 30778664 Free PMC article.
-
NADPH oxidases: Current aspects and tools.Redox Biol. 2020 Jul;34:101512. doi: 10.1016/j.redox.2020.101512. Epub 2020 May 23. Redox Biol. 2020. PMID: 32480354 Free PMC article. Review.
-
Increased Expression of DUOX2 Is an Epithelial Response to Mucosal Dysbiosis Required for Immune Homeostasis in Mouse Intestine.Gastroenterology. 2015 Dec;149(7):1849-59. doi: 10.1053/j.gastro.2015.07.062. Epub 2015 Aug 7. Gastroenterology. 2015. PMID: 26261005 Free PMC article.
References
-
- Dupuy C, Ohayon R, Valent A, Noël-Hudson MS, Dème D, Virion A. 1999. Purification of a novel flavoprotein involved in the thyroid NADPH oxidase. Cloning of the porcine and human cDNAs. J Biol Chem 274:37265–37269 - PubMed
-
- De Deken X, Wang D, Many MC, Costagliola S, Libert F, Vassart G, Dumont JE, Miot F. 2000. Cloning of two human thyroid cDNAs encoding new members of the NADPH oxidase family. J Biol Chem 275:23227–23233 - PubMed
-
- Rigutto S, Hoste C, Grasberger H, Milenkovic M, Communi D, Dumont JE, Corvilain B, Miot F, De Deken X. 2009. Activation of dual oxidases Duox1 and Duox2: differential regulation mediated by cAMP-dependent protein kinase and protein kinase c-dependent phosphorylation. J Biol Chem 284:6725–6734 - PMC - PubMed
-
- Corvilain B, van Sande J, Laurent E, Dumont JE. 1991. The H2O2-generating system modulates protein iodination and the activity of the pentose phosphate pathway in dog thyroid. Endocrinology 128:779–785 - PubMed
-
- Björkman U, Ekholm R. 1988. Accelerated exocytosis and H2O2 generation in isolated thyroid follicles enhance protein iodination. Endocrinology 122:488–494 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
