

doi: 10.4161/psb.7.1.18369.
Expression pattern and putative function of EXL1 and homologous genes in Arabidopsis
Affiliations
- PMID: 22301961
- PMCID: PMC3357360
- DOI: 10.4161/psb.7.1.18369
Item in Clipboard
Expression pattern and putative function of EXL1 and homologous genes in Arabidopsis
Plant Signal Behav.
2012 Jan.
Abstract
The Arabidopsis EXORDIUM-LIKE1 (EXL1) gene (At1g35140) is required for adaptation to carbon (C)- and energy-limiting growth conditions. An exl1 loss of function mutant showed diminished biomass production in a low total irradiance growth regime, impaired survival during extended night, and impaired survival of anoxia stress. We show here additional expression data and discuss the putative roles of EXL1. We hypothesize that EXL1 suppresses brassinosteroid-dependent growth and controls C allocation in the cell. In-depth expression analysis of homologous genes suggests that the EXL2 (At5g64260) and EXL4 (At5g09440) genes play similar roles.
Figures

Figure 1.EXL1, EXL2, and EXL4 expression in response to hypoxia. Ribosome-associated mRNAs were immunopurified from specific cell populations in the shoot, whole root, or root tip from 7-d-old Arabidopsis seedlings expressing a FLAG-epitope tagged ribosomal protein. Seedlings were cultured in the presence (control) or absence of air (2 h hypoxia). Affymetrix ATH1 CEL files (accession number GSE14578) were normalized and log2 transformed using RMA-Express. (A) Signal log ratios of relative transcript levels in different root tip cell types (hypoxia vs. control). (B) Signal log ratios of relative transcript levels in different root cell types (hypoxia vs. control). (C) Signal log ratios of relative transcript levels in different shoot cell types (hypoxia vs. control).
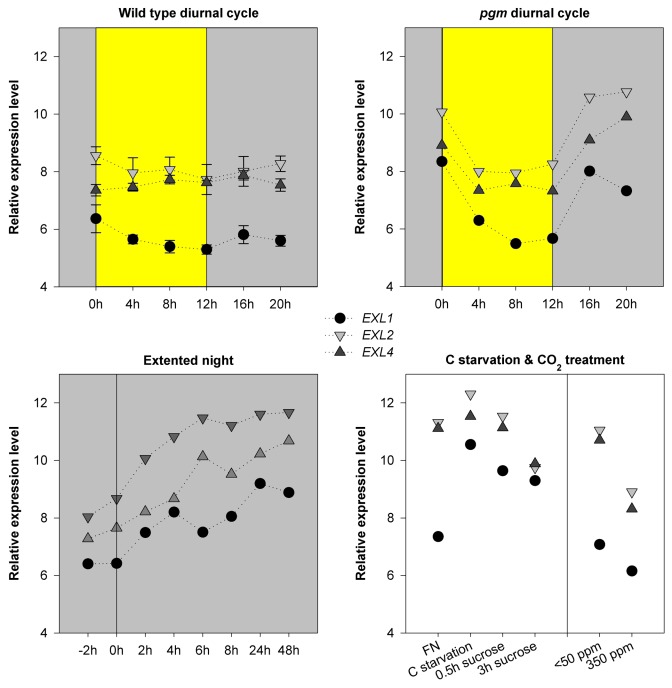
Figure 2.EXL1, EX2 and EXL4 expression in response to C availability. Publicly available expression profiles were normalized and log2 transformed using RMA-Express. The means of two or three replicates are shown. The standard error (SE) is shown if three replicates were available. Expression of EXL1 is shown for comparison. FN, full nutrition; C-starv, C starvation.

Figure 3. Quantitative RT-PCR analysis of EXL2 and EXL4 transcript levels in wild-type (Col-0) shoots. The mean of the CT (cycle threshold) values of three reference genes (eIF1α, PDF1, and LRS1) was subtracted from the respective CT value of the gene of interest. Subsequently, differences were subtracted from an arbitrary value (i.e., 40). Higher numbers indicate higher transcript levels. A difference of one unit indicates a fold change of approximately two. Error: SE of gene of interest in three technical replicates. eN: end of night; eD: end of day. (A) Plants were grown in soil under long-day conditions (16 h light/8 h night) and subjected to extended night. RNA was extracted from 28-d-old plants. (B) Plants were grown in soil under a low total irradiance regime [4 h light (60 µmol m−2 s−1)/20 h night]. RNA was extracted from shoots of 28-d-old plants.

Figure 4.EXL1, EXL2 and EXL4 expression in different organs and developmental stages. Wild-type expression profiles of the development series were downloaded from AtGenExpress and normalized and log2 transformed using RMA-Express., The mean and SD of three replicates are shown. Expression of EXL1 is shown for comparison.
References
-
- Gout E, Bligny R, Douce R, Boisson A-M, Rivasseu C. Early response of plant cell to carbon deprivation: in vivo 31P-NMR spectroscopy shows a quasi-instantaneous disruption on cytosolic sugars, phosphorylated intermediates of energy metabolism, phosphate partitioning, and intracellular pHs. New Phytol. 2011;189:135–47. doi: 10.1111/j.1469-8137.2010.03449.x. - DOI - PubMed
-
- Gibon Y, Bläsing OE, Palacios-Rojas N, Pankovic D, Hendriks JHM, Fisahn J, et al. Adjustment of diurnal starch turnover to short days: depletion of sugar during the night leads to a temporary inhibition of carbohydrate utilization, accumulation of sugars and post-translational activation of ADP-glucose pyrophosphorylase in the following light period. Plant J. 2004;39:847–62. doi: 10.1111/j.1365-313X.2004.02173.x. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases