

Curcumin suppresses T cell activation by blocking Ca2+ mobilization and nuclear factor of activated T cells (NFAT) activation
- PMID: 22303019
- PMCID: PMC3323045
- DOI: 10.1074/jbc.M111.318733
Curcumin suppresses T cell activation by blocking Ca2+ mobilization and nuclear factor of activated T cells (NFAT) activation
Abstract
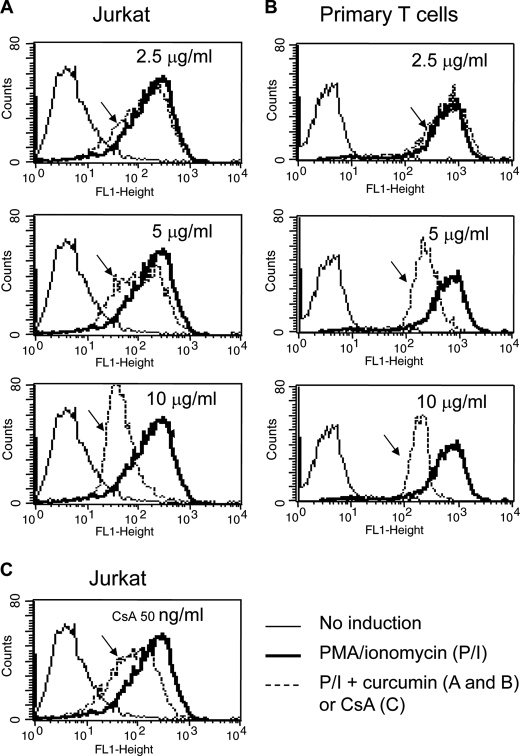
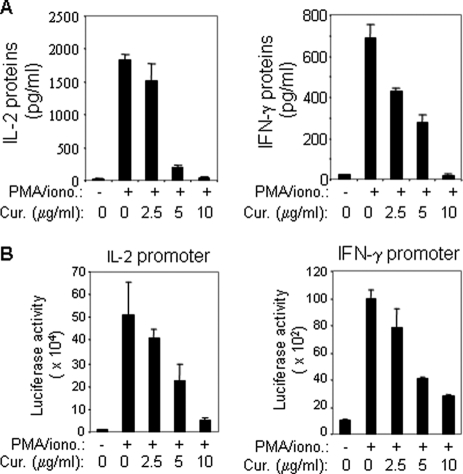
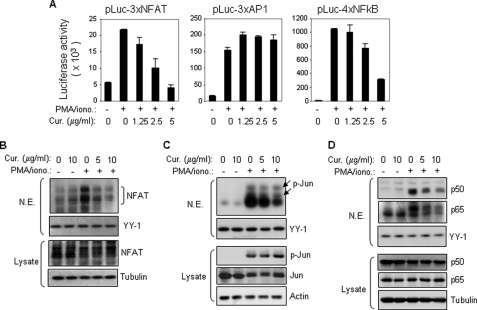
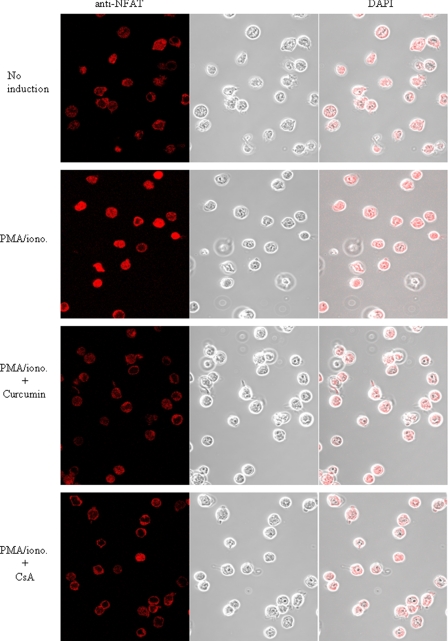
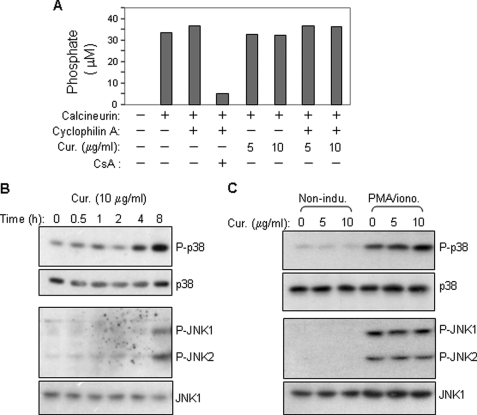
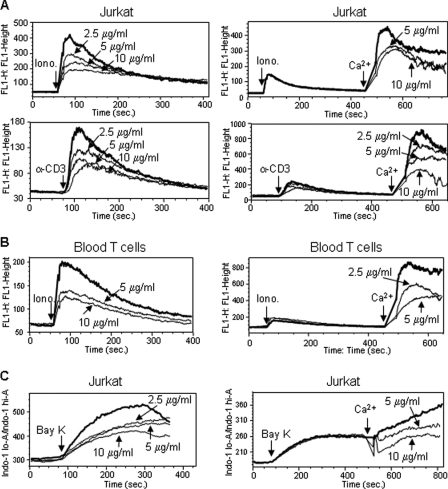
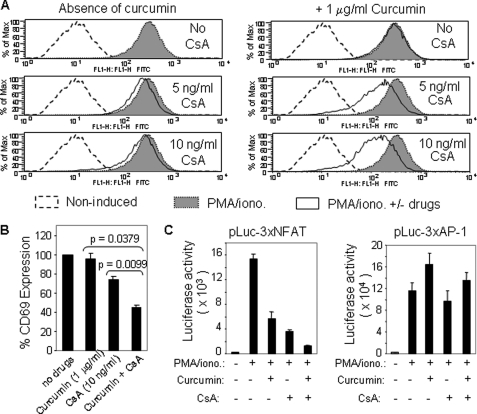
Curcumin is the active ingredient of the spice turmeric and has been shown to have a number of pharmacologic and therapeutic activities including antioxidant, anti-microbial, anti-inflammatory, and anti-carcinogenic properties. The anti-inflammatory effects of curcumin have primarily been attributed to its inhibitory effect on NF-κB activity due to redox regulation. In this study, we show that curcumin is an immunosuppressive phytochemical that blocks T cell-activation-induced Ca(2+) mobilization with IC(50) = ∼12.5 μM and thereby prevents NFAT activation and NFAT-regulated cytokine expression. This finding provides a new mechanism for curcumin-mediated anti-inflammatory and immunosuppressive function. We also show that curcumin can synergize with CsA to enhance immunosuppressive activity because of different inhibitory mechanisms. Furthermore, because Ca(2+) is also the secondary messenger crucial for the TCR-induced NF-κB signaling pathway, our finding also provides another mechanism by which curcumin suppresses NF-κB activation.
Figures
Similar articles
-
T Cell Receptor-induced Nuclear Factor κB (NF-κB) Signaling and Transcriptional Activation Are Regulated by STIM1- and Orai1-mediated Calcium Entry.J Biol Chem. 2016 Apr 15;291(16):8440-52. doi: 10.1074/jbc.M115.713008. Epub 2016 Jan 29. J Biol Chem. 2016. PMID: 26826124 Free PMC article.
-
Differential roles of PKC-theta in the regulation of intracellular calcium concentration in primary T cells.J Mol Biol. 2006 Jan 20;355(3):347-59. doi: 10.1016/j.jmb.2005.10.043. Epub 2005 Nov 8. J Mol Biol. 2006. PMID: 16309697
-
Human regulatory T cells rapidly suppress T cell receptor-induced Ca(2+), NF-κB, and NFAT signaling in conventional T cells.Sci Signal. 2011 Dec 20;4(204):ra90. doi: 10.1126/scisignal.2002179. Sci Signal. 2011. PMID: 22375050
-
Immunomodulation by curcumin.Adv Exp Med Biol. 2007;595:321-41. doi: 10.1007/978-0-387-46401-5_14. Adv Exp Med Biol. 2007. PMID: 17569218 Review.
-
Regulation of T-cell tolerance by calcium/NFAT signaling.Immunol Rev. 2009 Sep;231(1):225-40. doi: 10.1111/j.1600-065X.2009.00817.x. Immunol Rev. 2009. PMID: 19754900 Review.
Cited by
-
Curcuma Contra Cancer? Curcumin and Hodgkin's Lymphoma.Cancer Growth Metastasis. 2013 Aug 8;6:35-52. doi: 10.4137/CGM.S11113. eCollection 2013. Cancer Growth Metastasis. 2013. PMID: 24665206 Free PMC article. Review.
-
Hyaluronic acid-curcumin conjugate suppresses the fibrotic functions of myofibroblasts from contractive joint by the PTGER2 demethylation.Regen Biomater. 2019 Oct;6(5):269-277. doi: 10.1093/rb/rbz016. Epub 2019 Apr 22. Regen Biomater. 2019. PMID: 31616564 Free PMC article.
-
A Phase 2 Study to Assess the Immunomodulatory Capacity of a Lecithin-based Delivery System of Curcumin in Endometrial Cancer.Front Nutr. 2019 Jan 11;5:138. doi: 10.3389/fnut.2018.00138. eCollection 2018. Front Nutr. 2019. PMID: 30687714 Free PMC article.
-
Engineering Curcumin Biosynthesis in Poplar Affects Lignification and Biomass Yield.Front Plant Sci. 2022 Jul 4;13:943349. doi: 10.3389/fpls.2022.943349. eCollection 2022. Front Plant Sci. 2022. PMID: 35860528 Free PMC article.
-
The multifaceted role of curcumin in cancer prevention and treatment.Molecules. 2015 Feb 5;20(2):2728-69. doi: 10.3390/molecules20022728. Molecules. 2015. PMID: 25665066 Free PMC article. Review.
References
-
- Ammon H. P., Wahl M. A. (1991) Pharmacology of Curcuma longa. Planta Med. 57, 1–7 - PubMed
-
- Araújo C. C., Leon L. L. (2001) Biological activities of Curcuma longa L. Mem. Inst. Oswaldo. Cruz. 96, 723–728 - PubMed
-
- Jurenka J. S. (2009) Anti-inflammatory properties of curcumin, a major constituent of Curcuma longa: a review of preclinical and clinical research. Altern. Med. Rev. 14, 141–153 - PubMed
-
- Goel A., Kunnumakkara A. B., Aggarwal B. B. (2008) Curcumin as “Curecumin”: from kitchen to clinic. Biochem. Pharmacol. 75, 787–809 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
