

doi: 10.1199/tab.0129.
Epub 2010 Jan 22.
Jasmonates
Affiliations
- PMID: 22303255
- PMCID: PMC3244945
- DOI: 10.1199/tab.0129
Item in Clipboard
Jasmonates
Arabidopsis Book.
2010.
Abstract
ARABIDOPSIS IS A SUPERB MODEL FOR THE STUDY OF AN IMPORTANT SUBGROUP OF OXYLIPINS: the jasmonates. Jasmonates control many responses to cell damage and invasion and are essential for reproduction. Jasmonic acid (JA) is a prohormone and is conjugated to hydrophobic amino acids to produce regulatory ligands. The major receptor for active jasmonate ligands is closely related to auxin receptors and, as in auxin signaling, jasmonate signaling requires the destruction of repressor proteins. This chapter uses a frequently asked question (FAQ) approach and concludes with a practical section.
Figures
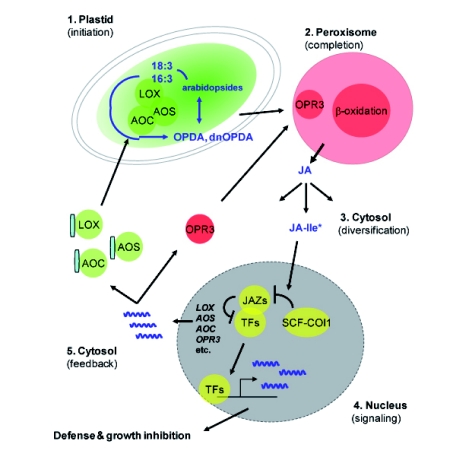
Cellular compartments in JA synthesis and signaling: an overview. The five major steps of JA synthesis and signaling (see text) take place in the several cellular compartments. JA is made by the combined action of plastids and peroxisomes. Many of the participating enzymes are jasmonate-inducible and targeted to these organelles. In the case of the plastid-localized enzymes LOX, AOS and AOC their import sequences (blue boxes) are cleaved off. Plastids also harbor pools of the JA precursors OPDA and dn-OPDA esterified to galactolipids in compounds known as arabidopsides. Jasmonoyl isoleucine (JA-Ile) made in the cytosol from JA functions as a signal in the nucleus. Note that several other compartments such as the cell walls and vacuoles might contribute to JA signaling (not shown).
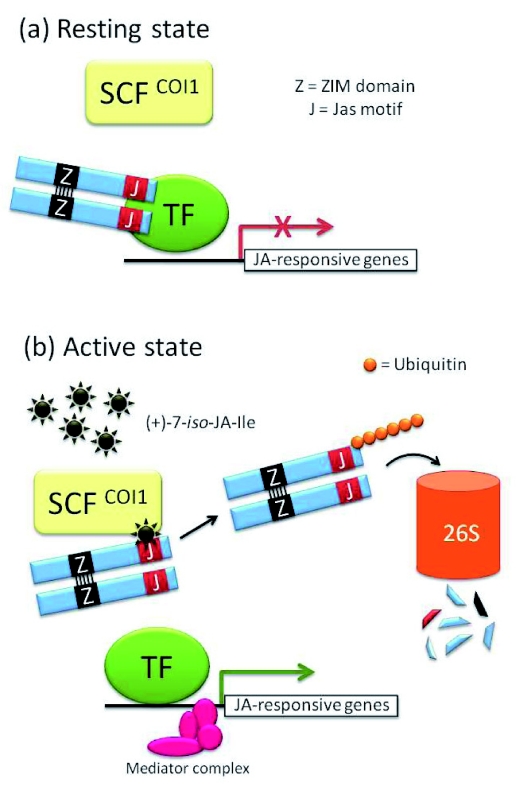
Model for jasmonate signaling In the resting state (a), when hormone concentrations are low or undetectable, JAZ proteins bind transcription factors such as MYC2 preventing their function as activators of jasmonate-responsive genes. In the active state (b), certain external stimuli (e.g. wounding) or developmental programs (e.g. anther maturation) promote jasmonate biosynthesis leading to active hormone accumulation (e.g. jasmonate isoleucine [(+)-7-iso-JA-Ile]). The hormone is perceived by the protein COI1 which is the F-box subunit of an E3 ubiquitin ligase of the type SKP1-CUL1-F-box (SCF). Hormone recognition by COI1 favors binding of COI1 to JAZ proteins via their Jas motif. In this model the JAZ proteins can be seen as co-receptors acting with COI1. This promotes ubiquitination of JAZ proteins and their subsequent degradation by the 26S proteasome. The transcription factors are now relieved from JAZ-mediated repression and free to recruit the RNA polymerase Il transcriptional machinery to the promoter of jasmonate-responsive genes. Recent evidence suggests that this recruitment occurs through universal adaptors such as the Mediator complex.
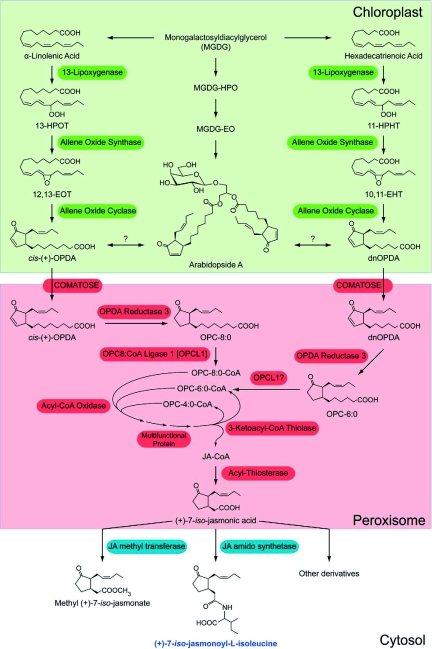
Jasmonic acid biosynthesis and metabolism in Arabidopsis Jasmonic acid (JA) is derived from polyunsaturated fatty acids linolenic acid (18:3) and hexadecatrienoic acid (16:3) which are stored in plastids mostly esterified to monogalactosyldiacylglycerol (MGDG). 13-LOXs add molecular oxygen to 18:3 and 16:3 to form their corresponding hydroperoxides 13(S)-hydroperoxy-octadecatrienoic acid [13-HPOT] and 11(S)-hydroperoxy-hexadecatrienoic acid [11-HPHT]. AOS transforms these into the allene oxides (13S)-12, 13-epoxy-octadecatrienoic acid [12, 13-EOT] and (11S)-10, 11-epoxy-octadecatrienoic acid [10, 11-EOT]. AOC directs the formation of a cyclopentenone ring in these compounds to yield 12-oxo-phytodienoic acid (OPDA) and dinor-oxo-phytodienoic acid (dnOPDA), respectively. OPDA and dnOPDA are also found esterified to galactolipids in the form of arabidopsides, which to date have only been isolated from Arabidopsis and a few other related crucifers. It is possible that these arabidopsides are synthesized through the direct action of 13-LOX, AOS and AOC in MGDG. OPDA and dnOPDA are transferred to the peroxisomes at least partially through the action of the ABC-type transporter COMATOSE (CTS). OPDA Reductase 3 catalyzes the reduction of the cyclopentenone ring in OPDA and dnOPDA to form 3-oxo-2-(2-(Z)-pentenyl)-cyclopentane-1-octanoic (OPC-8:0) and hexanoic (OPC-6:0) acids, respectively. The enzyme OPC-8:CoA ligase 1 (OPCL1) esterifies a CoA to the acyl group of OPC-8:0 and -presumably-OPC-6:0 in preparation for a series of ß-oxidation cycles that shorten the acidic chain of these compounds to yield jasmonic acid, ß-oxidation proceeds through three types of enzymes: acyl-CoA oxidase (ACX), multifunctional protein (MFP, which displays enoyl-CoA hydratase and ß-hydroxy-acyl-CoA dehydrogenase activities), and 3-ketoacyl-CoA thiolase (KAT). Finally, a presumed acyl-thioesterase (ACH) releases (+)-7-iso-jasmonic acid (JA) from its CoA ester. Upon transport to the cytoplasm, JA undergoes further metabolism such as esterification into methyl (+)-7-iso-jasmonate or amino acid conjugation to form (+)-7-iso-jasmonoyl-isoleucine which is considered the active form of the hormone. [Figure adapted from http://stke.sciencemag.org/cgi/cm/stkecm;CMP_7361 and Schaller and Stintzi (2009)]
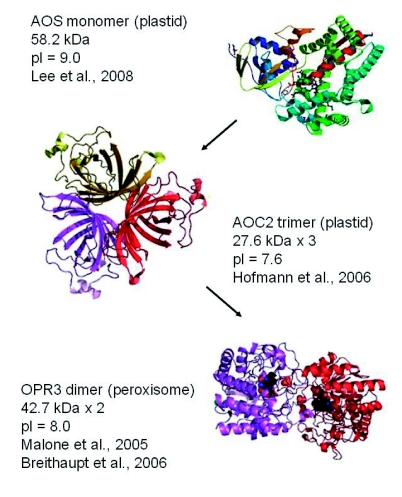
Structures of the three main enzymes of jasmonate synthesis Allene oxide synthase (AOS) is a basic protein that catalyses the formation of allene oxides from fatty acid hydroperoxides generated by lipoxygenases (LOXs). AOS crystallizes as a monomer. Allene oxide cyclase 2 (AOC2) is one of four AOCs encoded in the Arabidopsis genome. AOC2 directs the correct folding pathway of allene oxides into the natural stereochemical form of OPDA. The enzyme crystallizes as a trimer. Oxo-phytodienoic acid reductase 3 (OPR3) is an NADPH dehydrogenase that catalyses the reduction of the cyclopentenone double bond in OPDA to produce the intermediate OPC8. Tomato OPR3 crystallizes as a dimer (Breithaupt et al., 2006). Dimer formation was not observed in Arabidopsis OPR3 crystals (Malone et al., 2005) but based on Breithaupt et al. (2006) dimer formation is likely for the Arabidopsis enzyme and is thus indicated in the figure. The molecular mass and pl data given for the proteins includes the plastid import sequences present on AOS and AOC2.
References
-
- Bannenberg G., Martínez M., Ham berg M., Castresana C. Diversity of the enzymatic activity in the lipoxygenase gene family of Arabidopsis thaliana. Lipids. 2009a;448(1):85–95. - PubMed
-
- Bari R., Jones J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009;698(1):473–488. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials