

Phytochrome signaling mechanisms
- PMID: 22303272
- PMCID: PMC3268501
- DOI: 10.1199/tab.0148
Phytochrome signaling mechanisms
Abstract
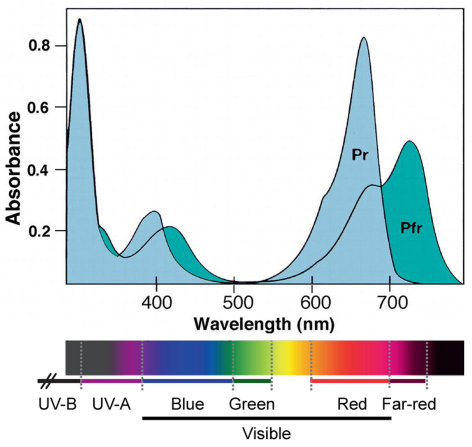
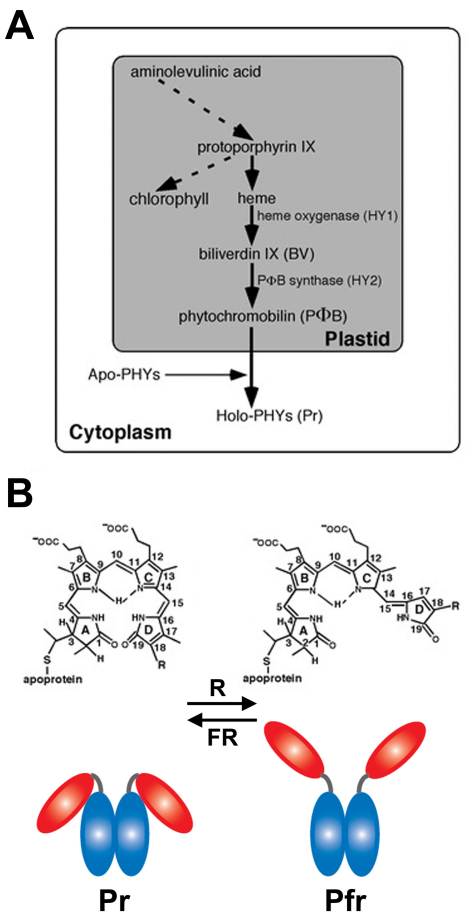
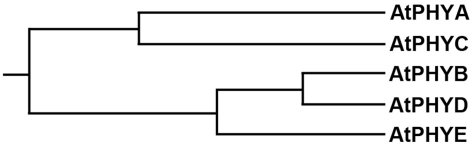
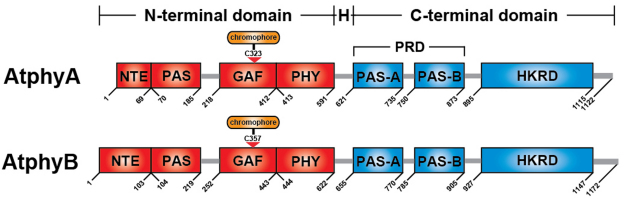
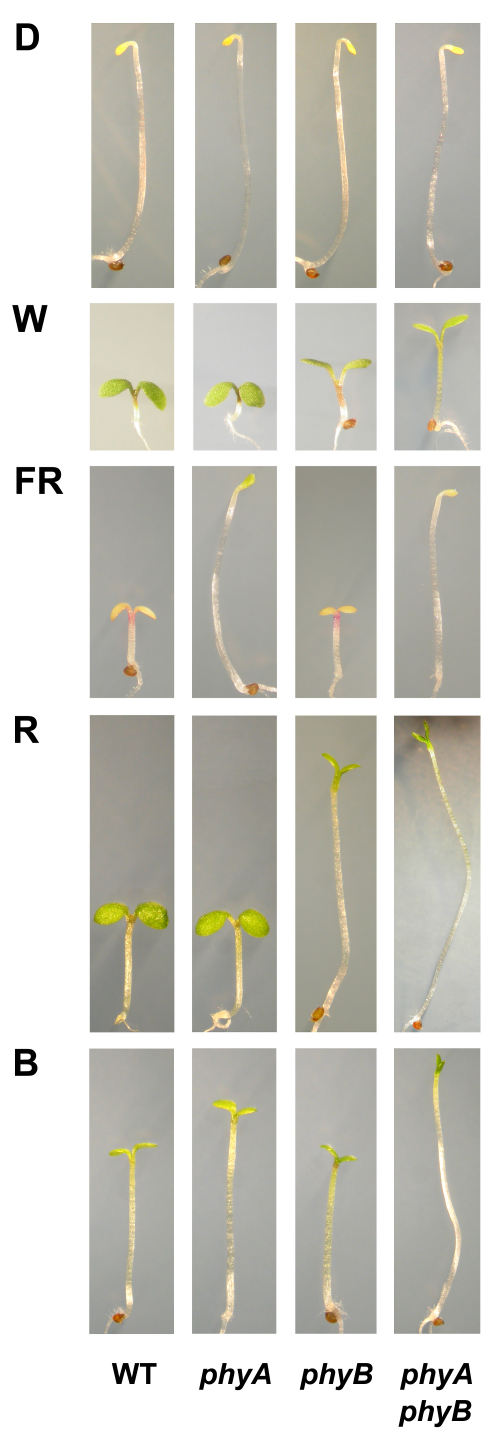
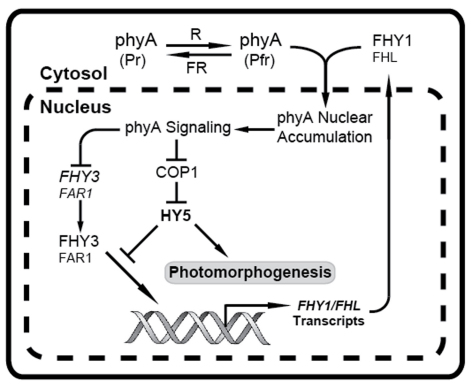
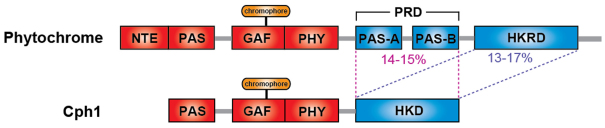
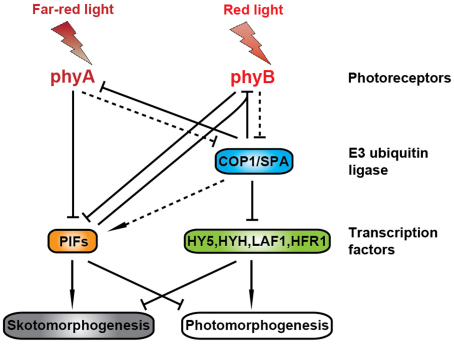
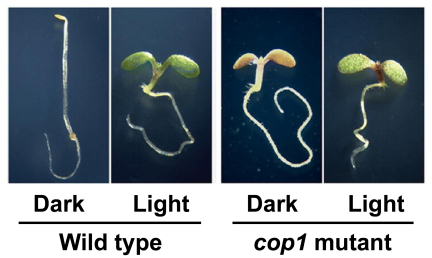
Phytochromes are red (R)/far-red (FR) light photoreceptors that play fundamental roles in photoperception of the light environment and the subsequent adaptation of plant growth and development. There are five distinct phytochromes in Arabidopsis thaliana, designated phytochrome A (phyA) to phyE. phyA is light-labile and is the primary photoreceptor responsible for mediating photomorphogenic responses in FR light, whereas phyB-phyE are light stable, and phyB is the predominant phytochrome regulating de-etiolation responses in R light. Phytochromes are synthesized in the cytosol in their inactive Pr form. Upon light irradiation, phytochromes are converted to the biologically active Pfr form, and translocate into the nucleus. phyB can enter the nucleus by itself in response to R light, whereas phyA nuclear import depends on two small plant-specific proteins FAR-RED ELONGATED HYPOCOTYL 1 (FHY1) and FHY1-LIKE (FHL). Phytochromes may function as light-regulated serine/threonine kinases, and can phosphorylate several substrates, including themselves in vitro. Phytochromes are phosphoproteins, and can be dephosphorylated by a few protein phosphatases. Photoactivated phytochromes rapidly change the expression of light-responsive genes by repressing the activity of CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1), an E3 ubiquitin ligase targeting several photomorphogenesis-promoting transcription factors for degradation, and by inducing rapid phosphorylation and degradation of Phytochrome-Interacting Factors (PIFs), a group of bHLH transcription factors repressing photomorphogenesis. Phytochromes are targeted by COP1 for degradation via the ubiquitin/26S proteasome pathway.
Figures

References
-
- Ahmad M., Cashmore A.R. The blue-light receptor cryptochrome 1 shows functional dependence on phytochrome A or phytochrome B in Arabidopsis thaliana. Plant J. 1997;11:421–427. - PubMed
-
- Ahmad M., Jarillo J.A., Smirnova O., Cashmore A.R. The CRY1 blue light photoreceptor of Arabidopsis interacts with phytochrome A in vitro. Mol. Cell. 1998;1:939–948. - PubMed
-
- Al-Sady B., Ni W., Kircher S., Schafer E., Quail P.H. Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Mol. Cell. 2006;23:439–446. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials