

Ret is a multifunctional coreceptor that integrates diffusible- and contact-axon guidance signals
- PMID: 22304922
- PMCID: PMC3286831
- DOI: 10.1016/j.cell.2012.01.024
Ret is a multifunctional coreceptor that integrates diffusible- and contact-axon guidance signals
Abstract
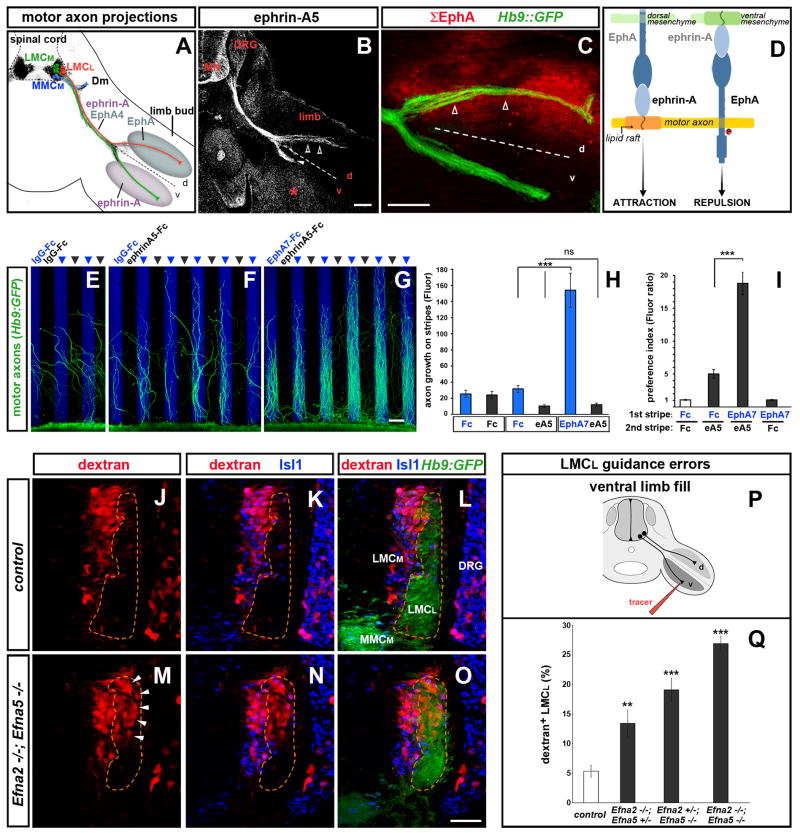
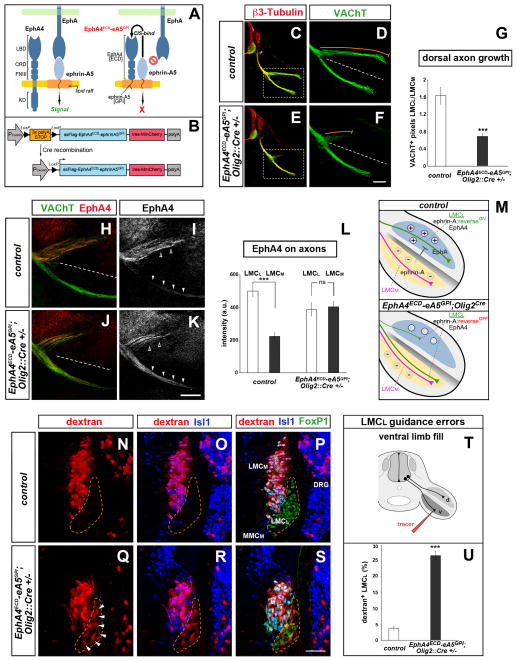
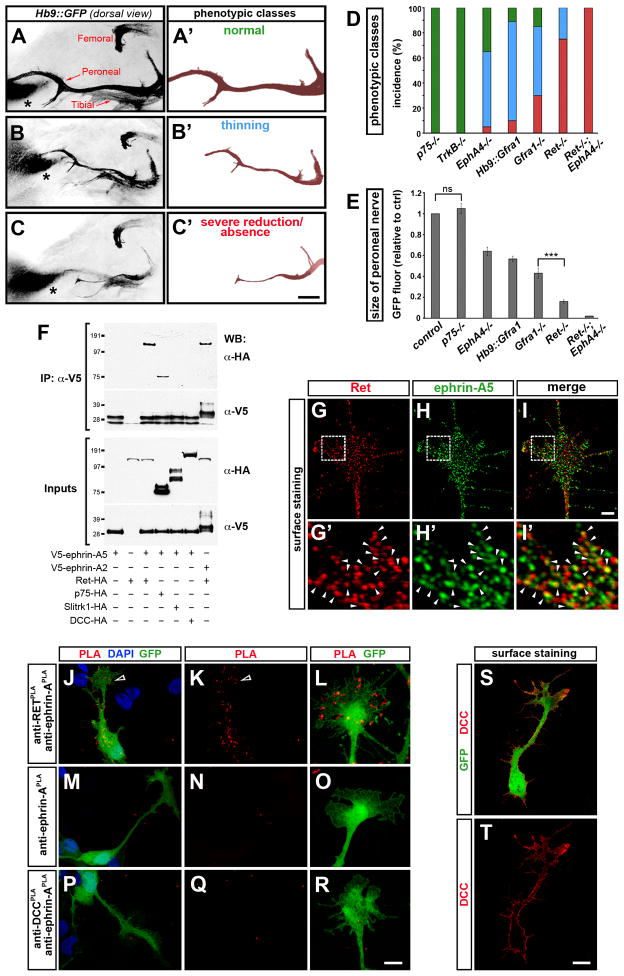
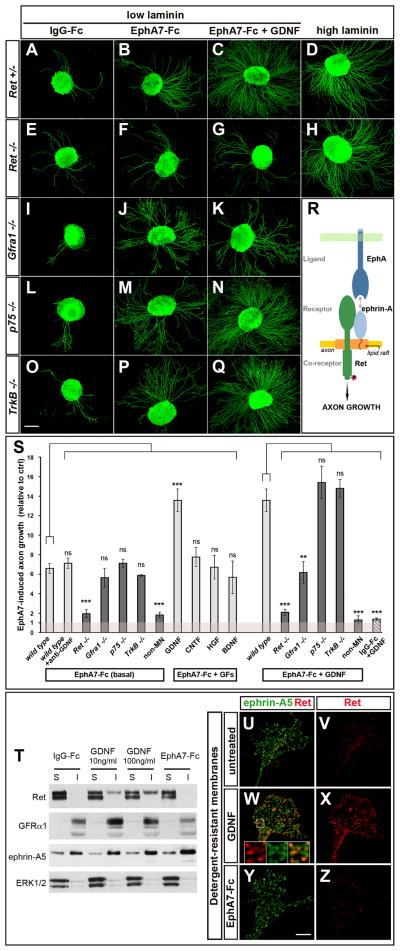
Growing axons encounter multiple guidance cues, but it is unclear how separate signals are resolved and integrated into coherent instructions for growth cone navigation. We report that glycosylphosphatidylinositol (GPI)-anchored ephrin-As function as "reverse" signaling receptors for motor axons when contacted by transmembrane EphAs present in the dorsal limb. Ephrin-A receptors are thought to depend on transmembrane coreceptors for transmitting signals intracellularly. We show that the receptor tyrosine kinase Ret is required for motor axon attraction mediated by ephrin-A reverse signaling. Ret also mediates GPI-anchored GFRα1 signaling in response to GDNF, a diffusible chemoattractant in the limb, indicating that Ret is a multifunctional coreceptor for guidance molecules. Axons respond synergistically to coactivation by GDNF and EphA ligands, and these cooperative interactions are gated by GFRα1 levels. Our studies uncover a hierarchical GPI-receptor signaling network that is constructed from combinatorial components and integrated through Ret using ligand coincidence detection.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
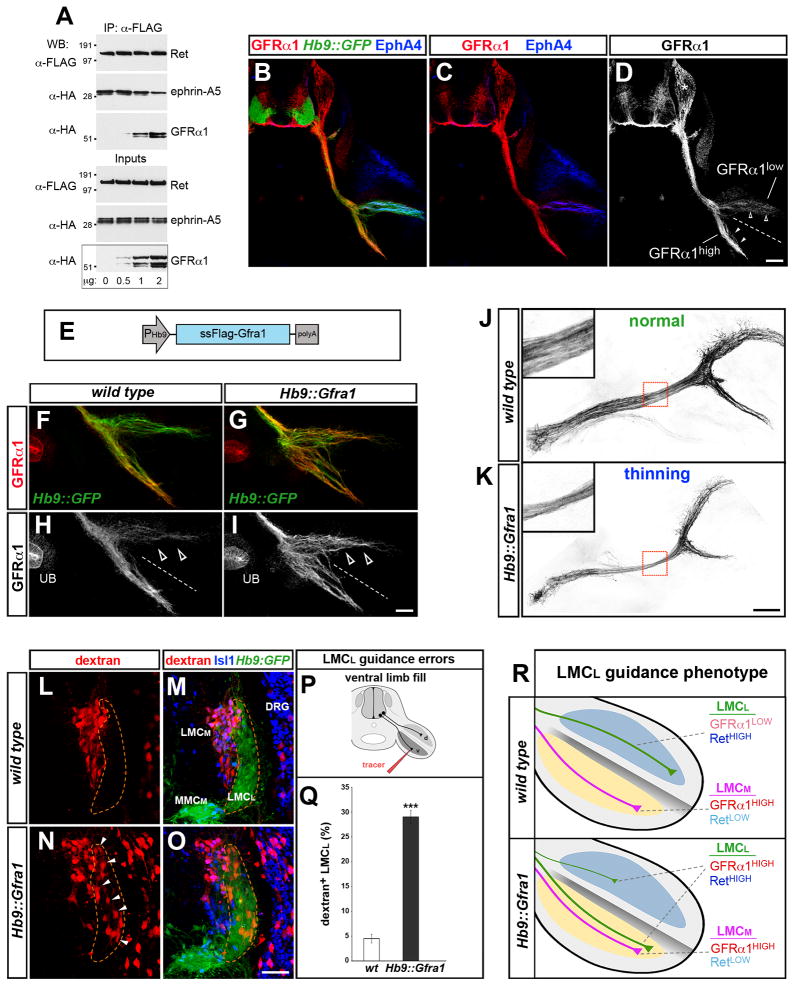
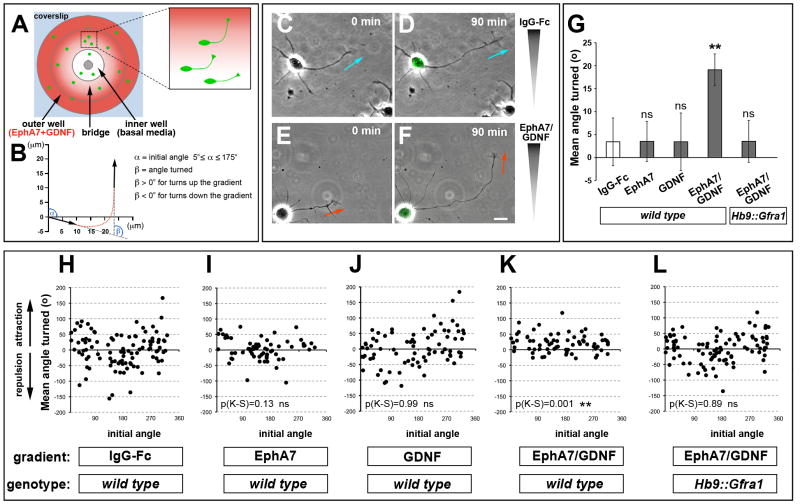
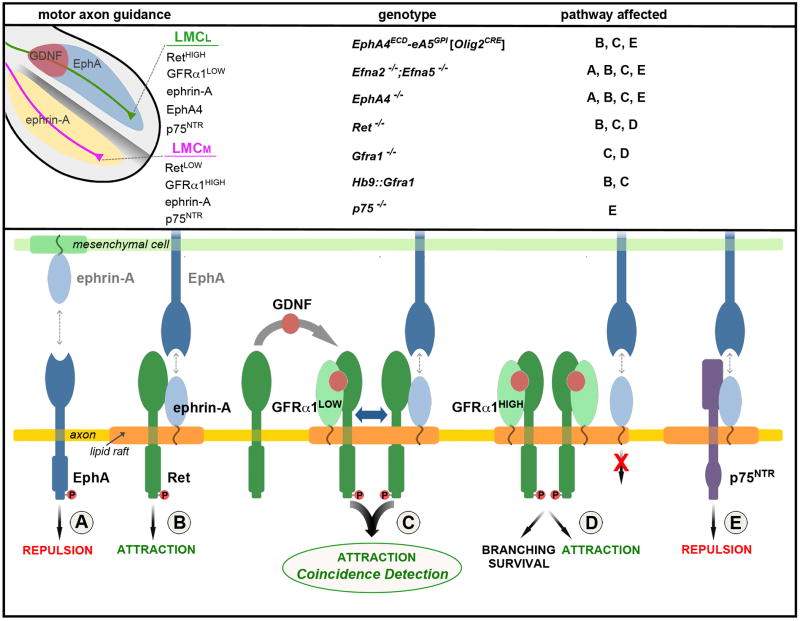
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
