

Regulation of circadian behavioral output via a MicroRNA-JAK/STAT circuit
- PMID: 22305007
- PMCID: PMC3307393
- DOI: 10.1016/j.cell.2011.12.024
Regulation of circadian behavioral output via a MicroRNA-JAK/STAT circuit
Abstract
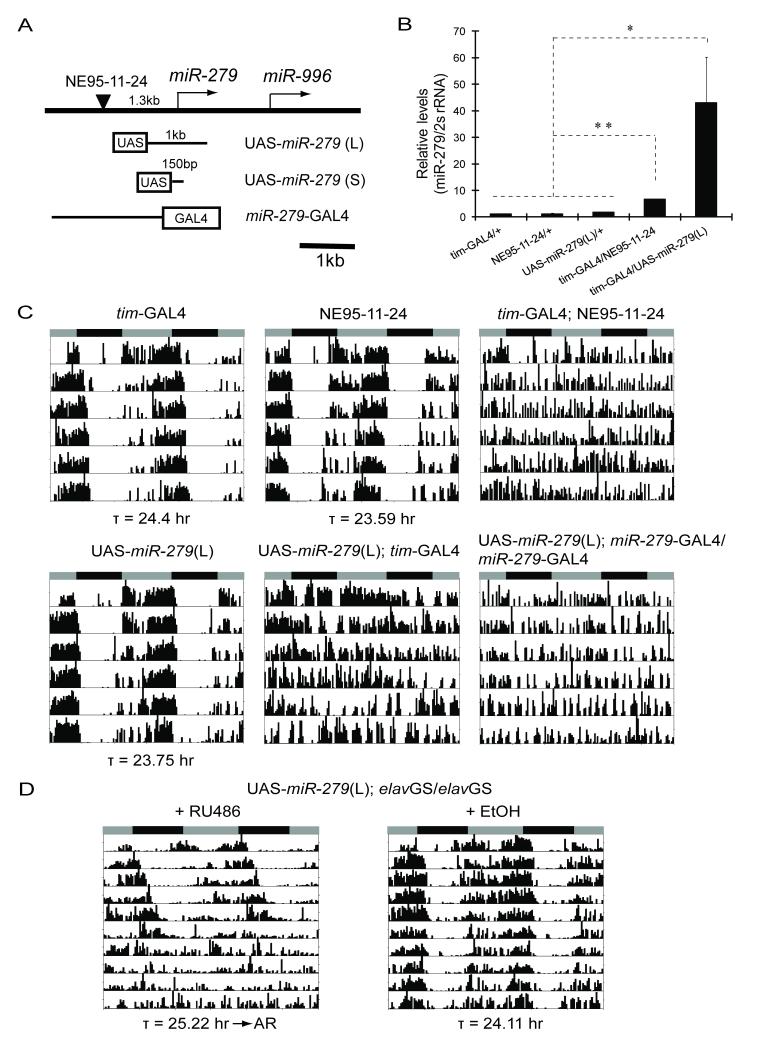
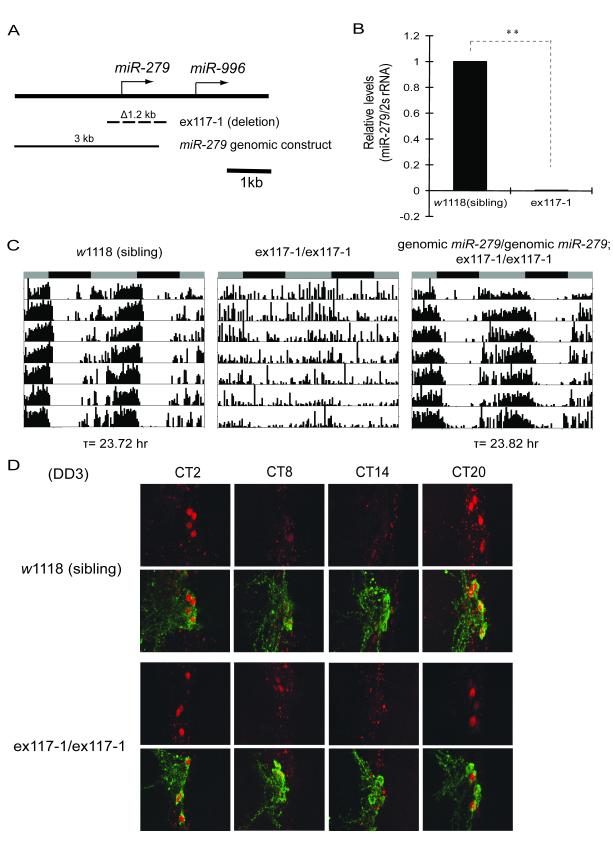
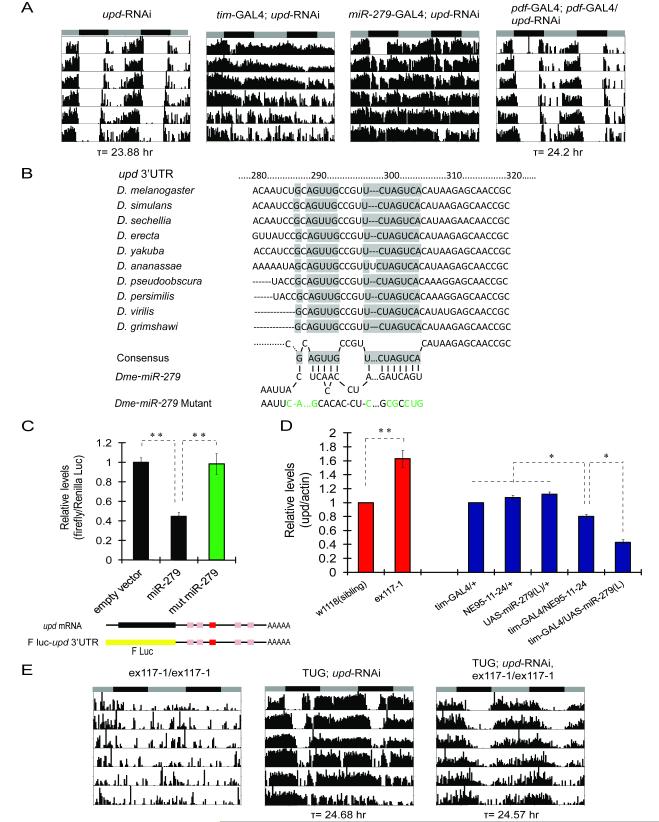
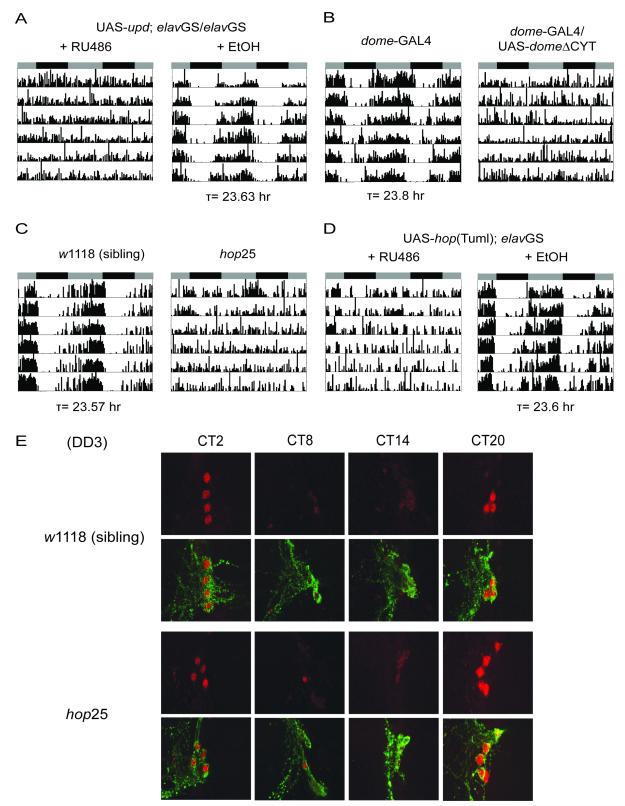
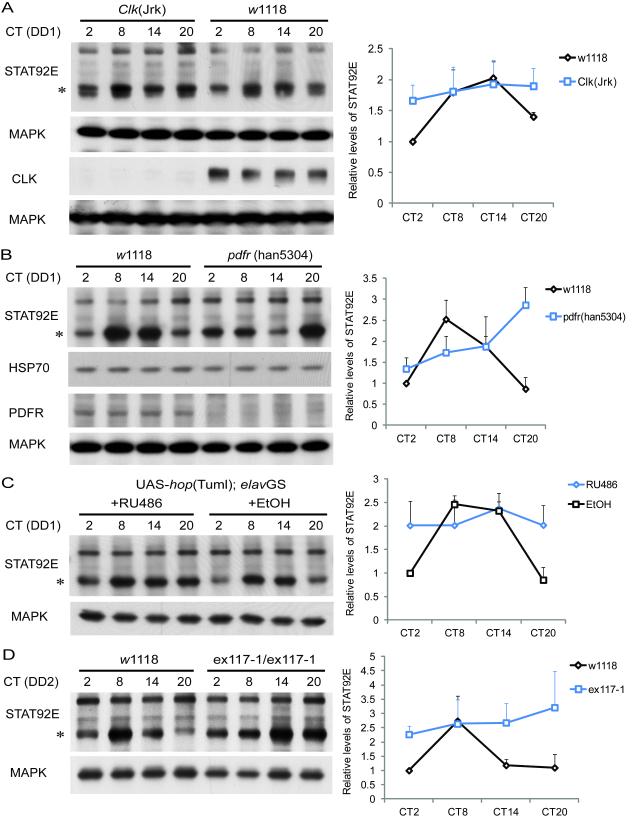
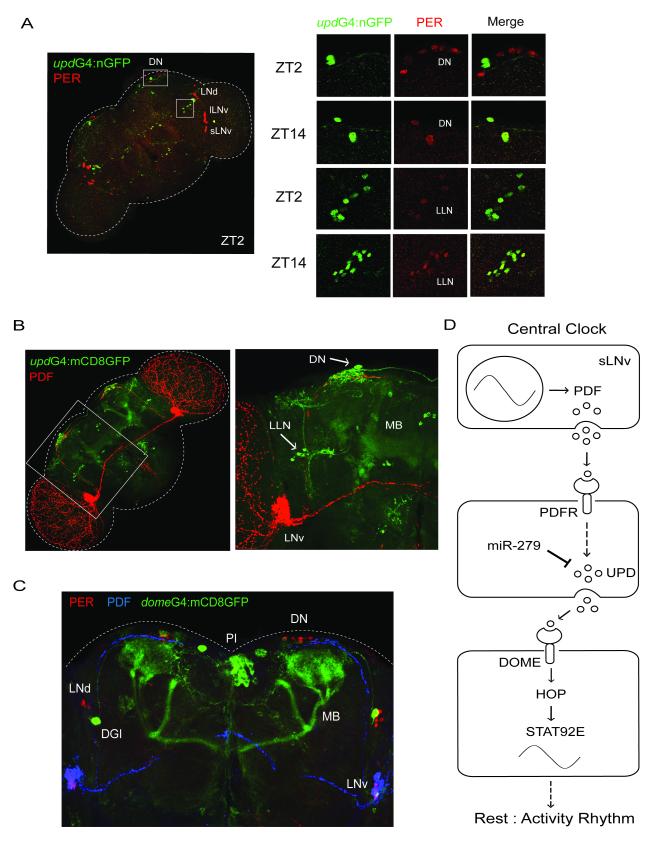
Although molecular components of the circadian clock are known, mechanisms that transmit signals from the clock and produce rhythmic behavior are poorly understood. We find that the microRNA miR-279 regulates the JAK/STAT pathway to drive rest:activity rhythms in Drosophila. Overexpression of microRNA miR-279 or miR-279 deletion attenuates rest:activity rhythms. Oscillations of the clock protein PERIOD are normal in pacemaker neurons lacking miR-279, suggesting that miR-279 acts downstream of the clock. We identify the JAK/STAT ligand, Upd, as a target of miR-279 and show that knockdown of Upd rescues the behavioral phenotype of miR-279 mutants. Manipulations of the JAK/STAT pathway also disrupt circadian rhythms. In addition, central clock neurons project in the vicinity of Upd-expressing neurons, providing a possible physical connection by which the central clock could regulate JAK/STAT signaling to control rest:activity rhythms.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Arbouzova NI, Zeidler MP. JAK/STAT signalling in Drosophila: insights into conserved regulatory and cellular functions. Development. 2006;133:2605–2616. - PubMed
-
- Asher G, Schibler U. Crosstalk between components of circadian and metabolic cycles in mammals. Cell Metab. 2011;13:125–137. - PubMed
-
- Banerjee S, Byrd JN, Gianino SM, Harpstrite SE, Rodriguez FJ, Tuskan RG, Reilly KM, Piwnica-Worms DR, Gutmann DH. The neurofibromatosis type 1 tumor suppressor controls cell growth by regulating signal transducer and activator of transcription-3 activity in vitro and in vivo. Cancer Res. 2010;70:1356–1366. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
