

Interaction of J-protein co-chaperone Jac1 with Fe-S scaffold Isu is indispensable in vivo and conserved in evolution
- PMID: 22306468
- PMCID: PMC3288383
- DOI: 10.1016/j.jmb.2012.01.022
Interaction of J-protein co-chaperone Jac1 with Fe-S scaffold Isu is indispensable in vivo and conserved in evolution
Abstract
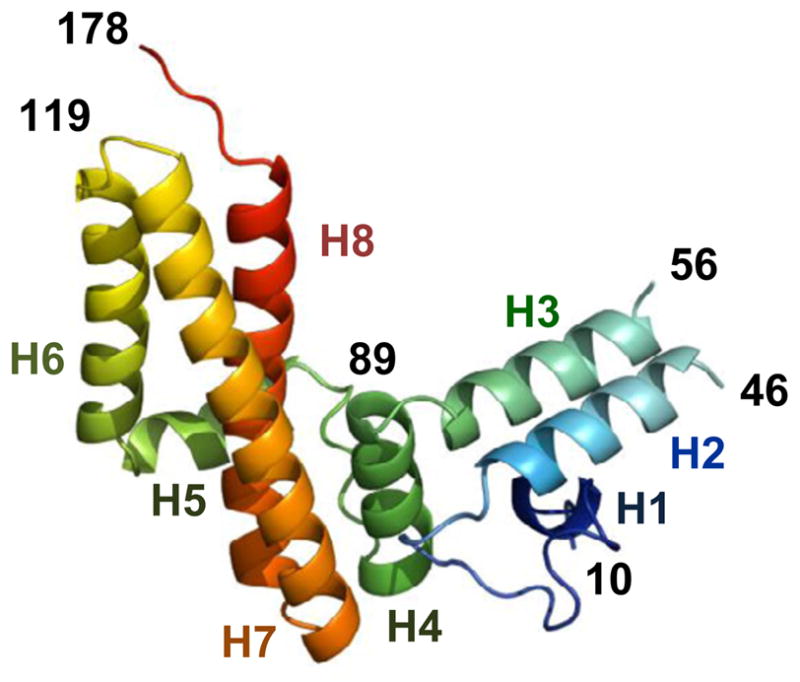
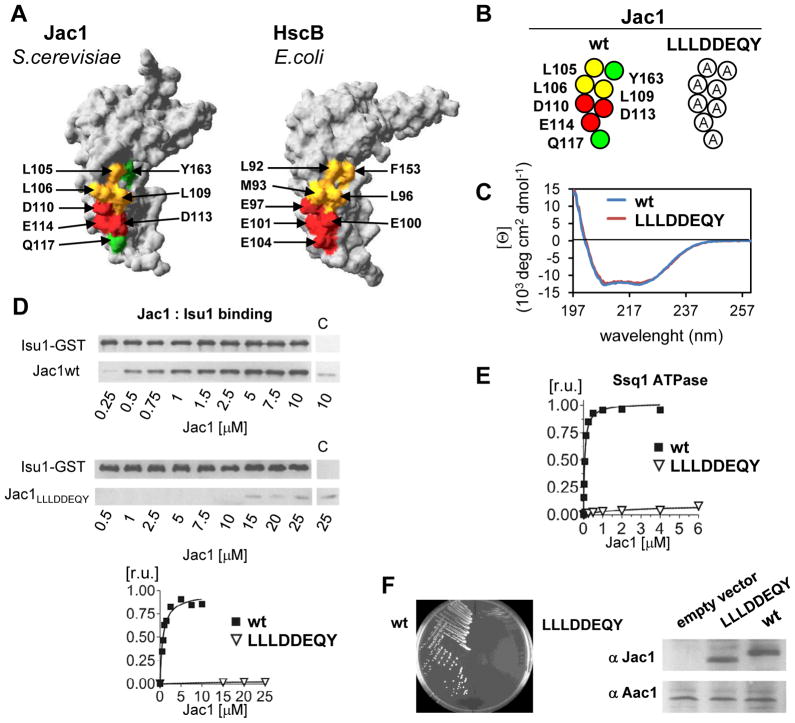
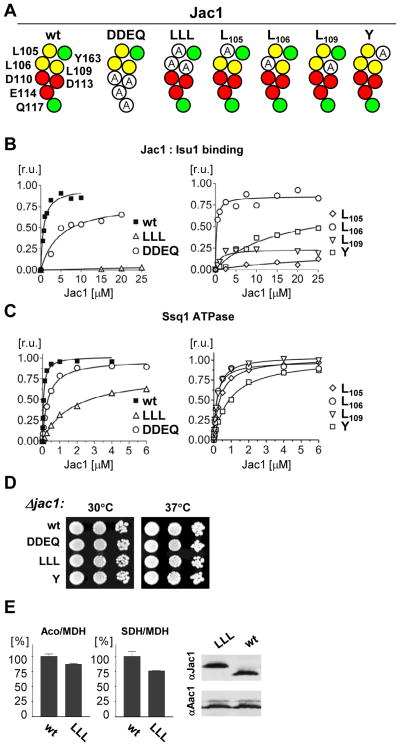
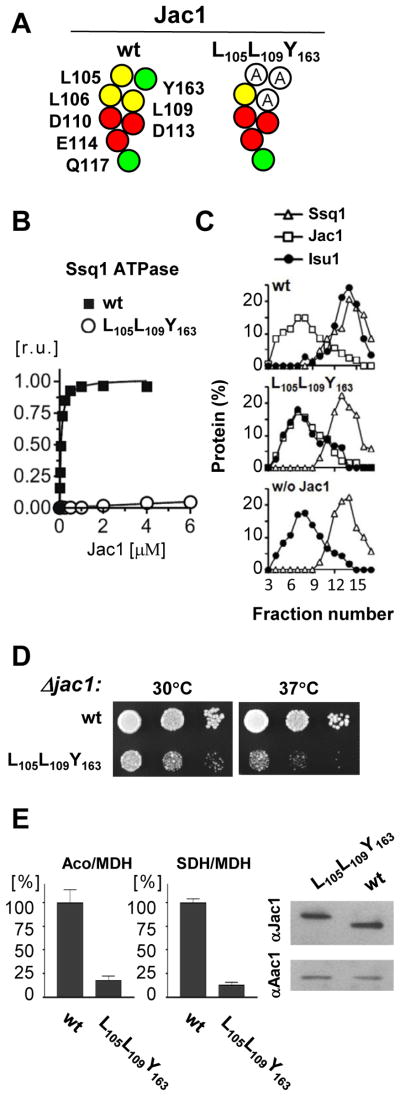
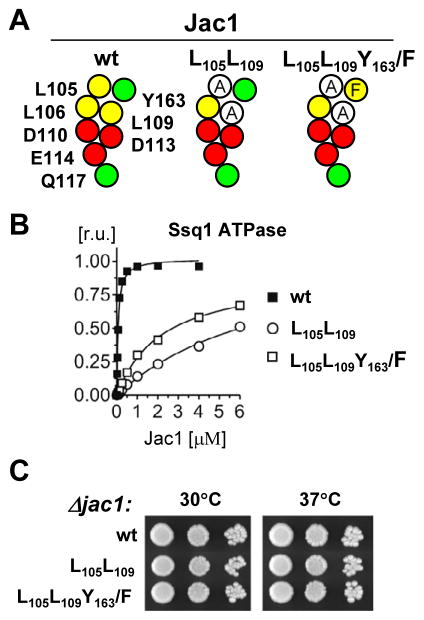
The ubiquitous mitochondrial J-protein Jac1, called HscB in Escherichia coli, and its partner Hsp70 play a critical role in the transfer of Fe-S clusters from the scaffold protein Isu to recipient proteins. Biochemical results from eukaryotic and prokaryotic systems indicate that formation of the Jac1-Isu complex is important for both targeting of the Isu for Hsp70 binding and stimulation of Hsp70's ATPase activity. However, in apparent contradiction, we previously reported that an 8-fold decrease in Jac1's affinity for Isu1 is well tolerated in vivo, raising the question as to whether the Jac1:Isu interaction actually plays an important biological role. Here, we report the determination of the structure of Jac1 from Saccharomyces cerevisiae. Taking advantage of this information and recently published data from the homologous bacterial system, we determined that a total of eight surface-exposed residues play a role in Isu binding, as assessed by a set of biochemical assays. A variant having alanines substituted for these eight residues was unable to support growth of a jac1-Δ strain. However, replacement of three residues caused partial loss of function, resulting in a significant decrease in the Jac1:Isu1 interaction, a slow growth phenotype, and a reduction in the activity of Fe-S cluster-containing enzymes. Thus, we conclude that the Jac1:Isu1 interaction plays an indispensable role in the essential process of mitochondrial Fe-S cluster biogenesis.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Lill R, Muhlenhoff U. Maturation of iron-sulfur proteins in eukaryotes: mechanisms, connected processes, and diseases. Annu Rev Biochem. 2008;77:669–700. - PubMed
-
- Garland SA, Hoff K, Vickery LE, Culotta VC. Saccharomyces cerevisiae ISU1 and ISU2: members of a well-conserved gene family for iron-sulfur cluster assembly. J Mol Biol. 1999;294:897–907. - PubMed
-
- Tsai CL, Barondeau DP. Human frataxin is an allosteric switch that activates the FeS cluster biosynthetic complex. Biochemistry. 2010;49:9132–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
