

The glutamate transporter, GLAST, participates in a macromolecular complex that supports glutamate metabolism
- PMID: 22306776
- PMCID: PMC3350823
- DOI: 10.1016/j.neuint.2012.01.013
The glutamate transporter, GLAST, participates in a macromolecular complex that supports glutamate metabolism
Abstract
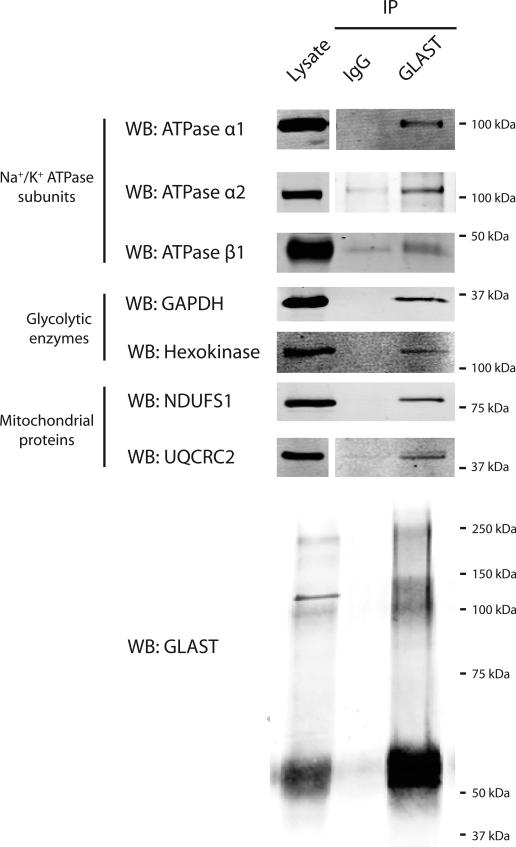
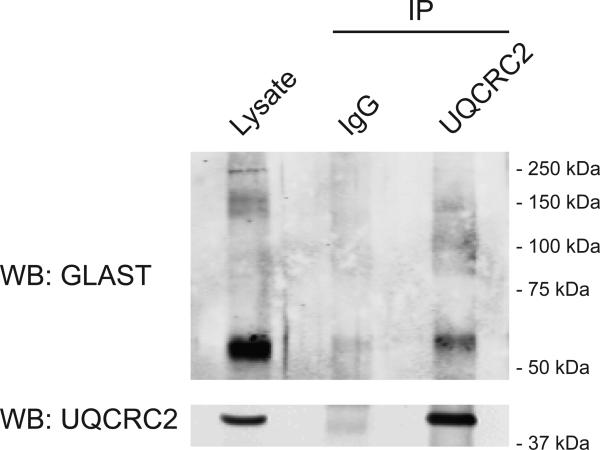
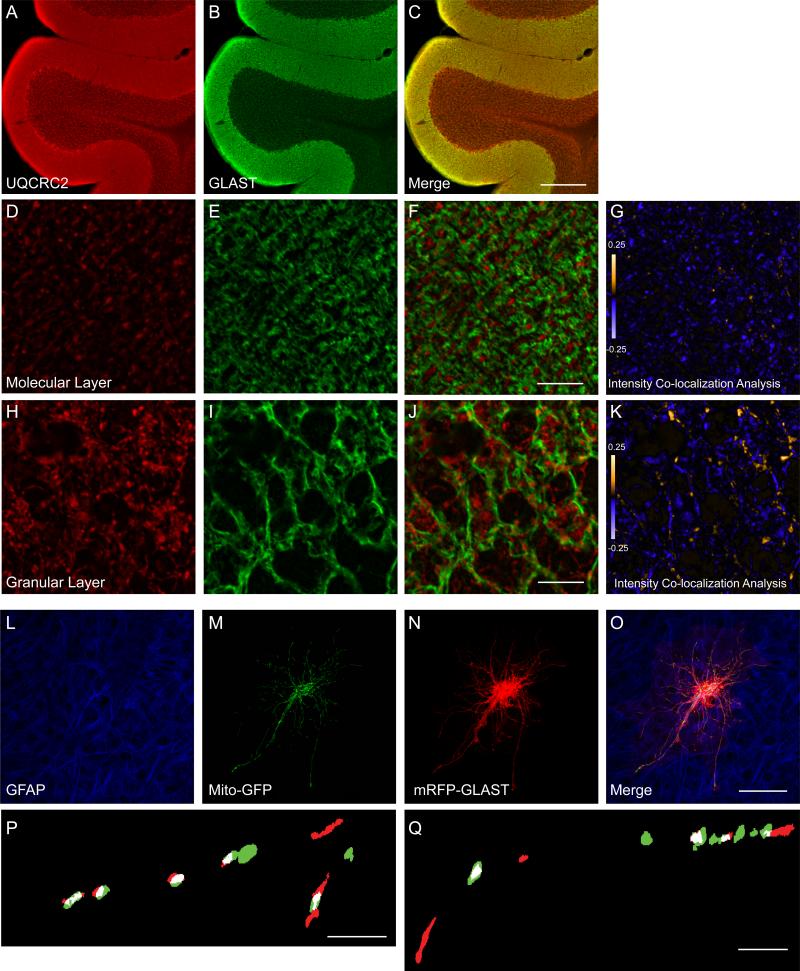
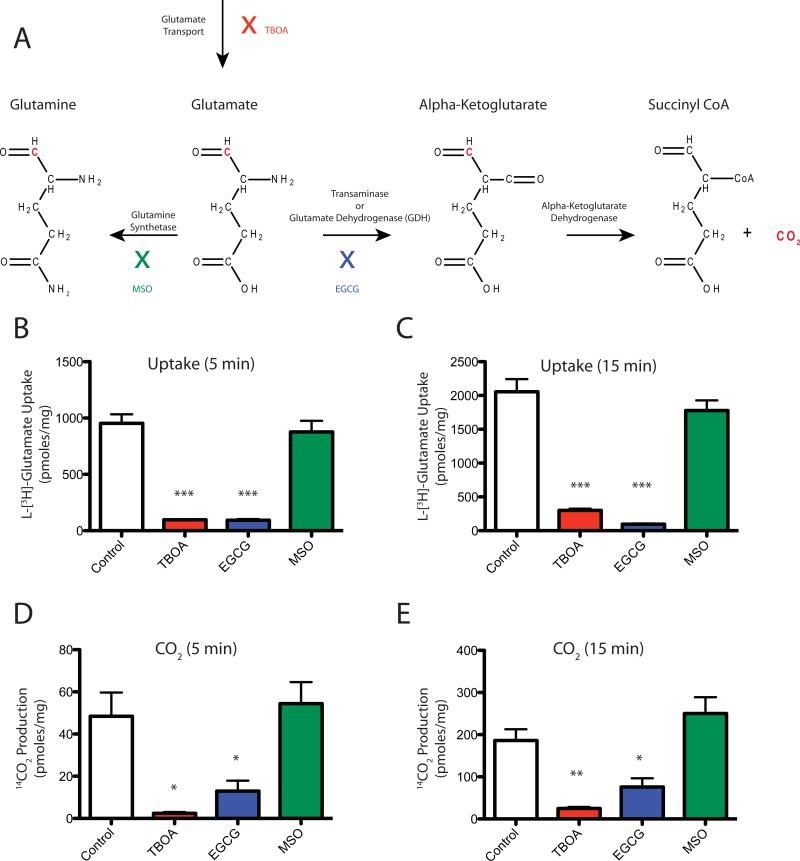
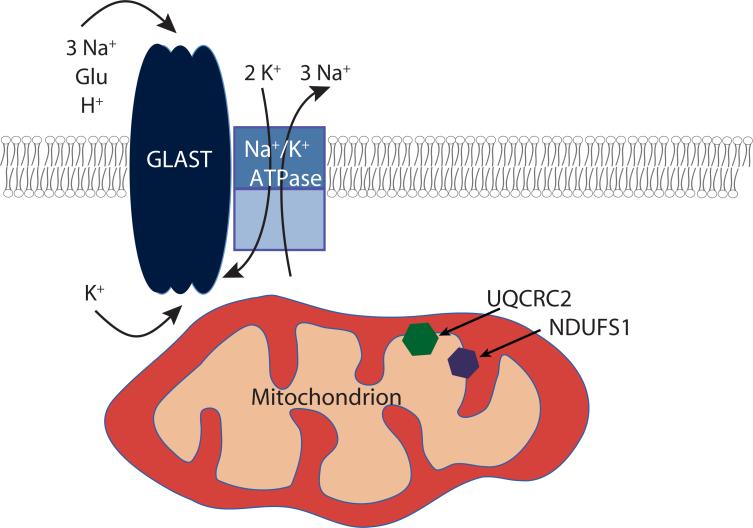
GLAST is the predominant glutamate transporter in the cerebellum and contributes substantially to glutamate transport in forebrain. This astroglial glutamate transporter quickly binds and clears synaptically released glutamate and is principally responsible for ensuring that synaptic glutamate concentrations remain low. This process is associated with a significant energetic cost. Compartmentalization of GLAST with mitochondria and proteins involved in energy metabolism could provide energetic support for glutamate transport. Therefore, we performed immunoprecipitation and co-localization experiments to determine if GLAST might co-compartmentalize with proteins involved in energy metabolism. GLAST was immunoprecipitated from rat cerebellum and subunits of the Na(+)/K(+) ATPase, glycolytic enzymes, and mitochondrial proteins were detected. GLAST co-localized with mitochondria in cerebellar tissue. GLAST also co-localized with mitochondria in fine processes of astrocytes in organotypic hippocampal slice cultures. From these data, we hypothesized that mitochondria participate in a macromolecular complex with GLAST to support oxidative metabolism of transported glutamate. To determine the functional metabolic role of this complex, we measured CO(2) production from radiolabeled glutamate in cultured astrocytes and compared it to overall glutamate uptake. Within 15 min, 9% of transported glutamate was converted to CO(2). This CO(2) production was blocked by inhibitors of glutamate transport and glutamate dehydrogenase, but not by an inhibitor of glutamine synthetase. Our data support a model in which GLAST exists in a macromolecular complex that allows transported glutamate to be metabolized in mitochondria to support energy production.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures

References
-
- Abu-Hamad S, Zaid H, Israelson A, Nahon E, Shoshan-Barmatz V. Hexokinase-I protection against apoptotic cell death is mediated via interaction with the voltage-dependent anion channel-1: mapping the site of binding. J. Biol. Chem. 2008;283:13482–13490. - PubMed
-
- Benediktsson AM, Schachtele SJ, Green SH, Dailey ME. Ballistic labeling and dynamic imaging of astrocytes in organotypic hippocampal slice cultures. J Neurosci Methods. 2005;141:41–53. - PubMed
-
- Bernardinelli Y, Azarias G, Chatton JY. In situ fluorescence imaging of glutamate-evoked mitochondrial Na+ responses in astrocytes. Glia. 2006;54:460–470. - PubMed
-
- Beutner G, Ruck A, Riede B, Brdiczka D. Complexes between porin, hexokinase, mitochondrial creatine kinase and adenylate translocator display properties of the permeability transition pore. Implication for regulation of permeability transition by the kinases. Biochim. Biophys. Acta. 1998;1368:7–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
