

Coordination of multiple enzyme activities by a single PCNA in archaeal Okazaki fragment maturation
- PMID: 22307085
- PMCID: PMC3321178
- DOI: 10.1038/emboj.2012.12
Coordination of multiple enzyme activities by a single PCNA in archaeal Okazaki fragment maturation
Abstract
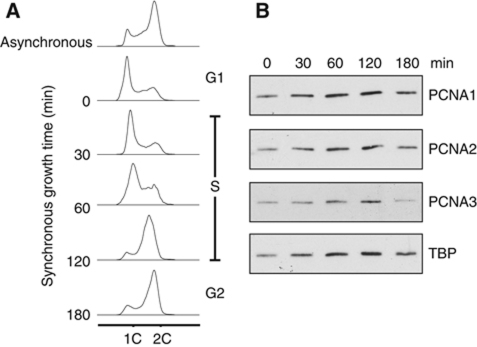
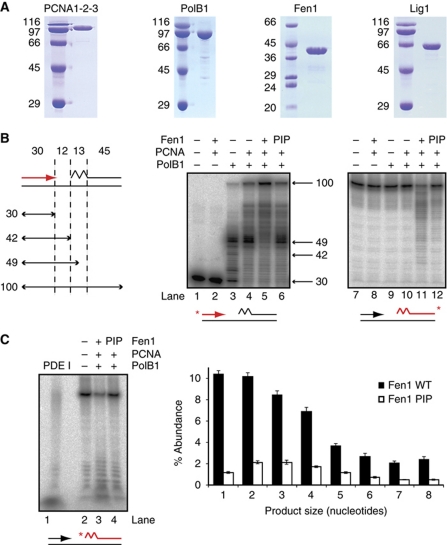
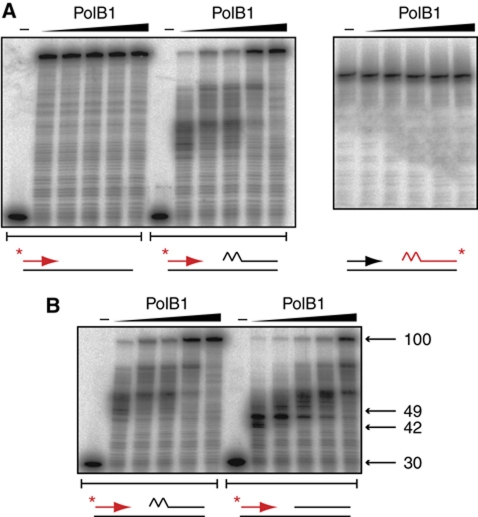
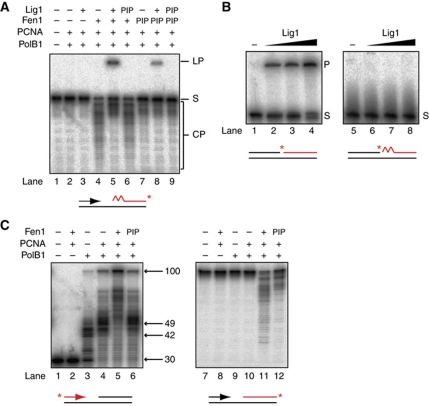
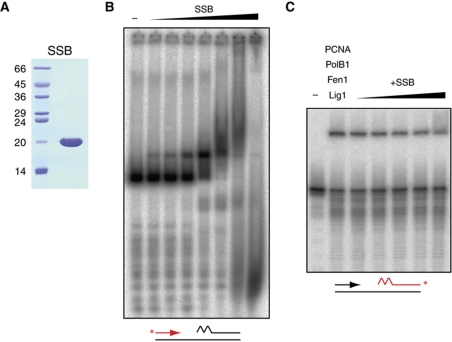
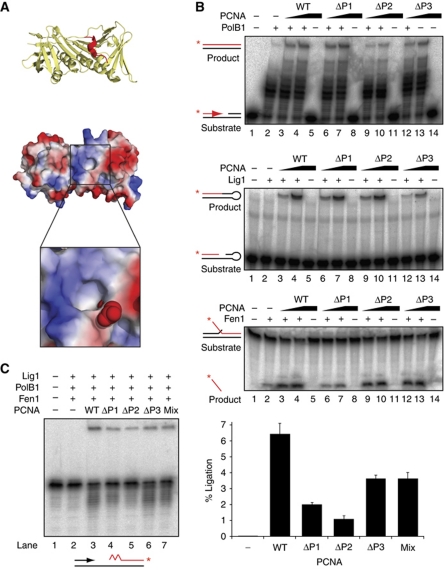
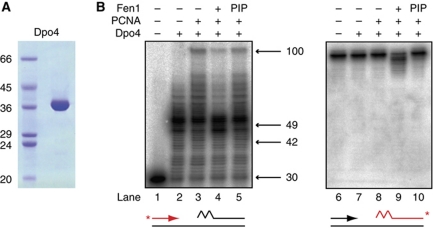
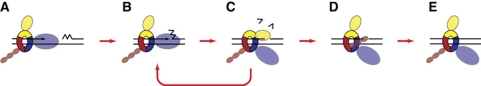
Chromosomal DNA replication requires one daughter strand-the lagging strand-to be synthesised as a series of discontinuous, RNA-primed Okazaki fragments, which must subsequently be matured into a single covalent DNA strand. Here, we describe the reconstitution of Okazaki fragment maturation in vitro using proteins derived from the archaeon Sulfolobus solfataricus. Six proteins are necessary and sufficient for coupled DNA synthesis, RNA primer removal and DNA ligation. PolB1, Fen1 and Lig1 provide the required catalytic activities, with coordination of their activities dependent upon the DNA sliding clamp, proliferating cell nuclear antigen (PCNA). S. solfataricus PCNA is a heterotrimer, with each subunit having a distinct specificity for binding PolB1, Fen1 or Lig1. Our data demonstrate that the most efficient coupling of activities occurs when a single PCNA ring organises PolB1, Fen1 and Lig1 into a complex.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Ayyagari R, Gomes XV, Gordenin DA, Burgers PM (2003) Okazaki fragment maturation in yeast. I. Distribution of functions between FEN1 AND DNA2. J Biol Chem 278: 1618–1625 - PubMed
-
- Beattie TR, Bell SD (2011a) The role of the DNA sliding clamp in Okazaki fragment maturation in archaea and eukaryotes. Biochem Soc Trans 39: 70–76 - PubMed
-
- Beattie TR, Bell SD (2011b) Molecular machines in archaeal DNA replication. Curr Opin Chem Biol 15: 614–619 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
