

Rerouting carbon flux to enhance photosynthetic productivity
- PMID: 22307292
- PMCID: PMC3318813
- DOI: 10.1128/AEM.07901-11
Rerouting carbon flux to enhance photosynthetic productivity
Abstract
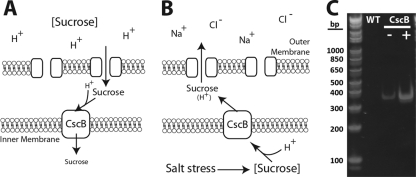
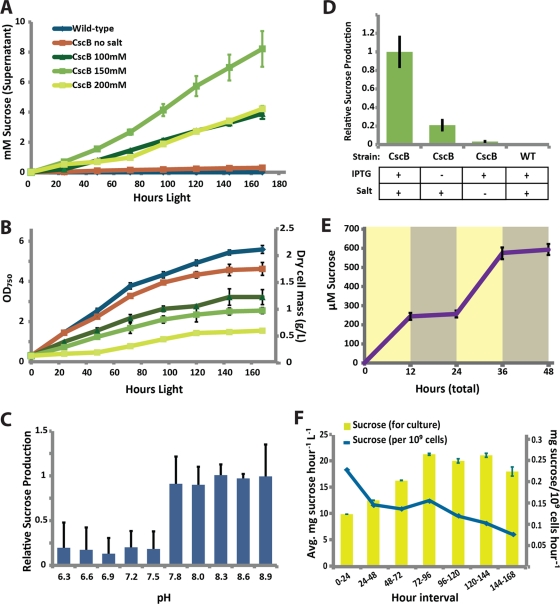
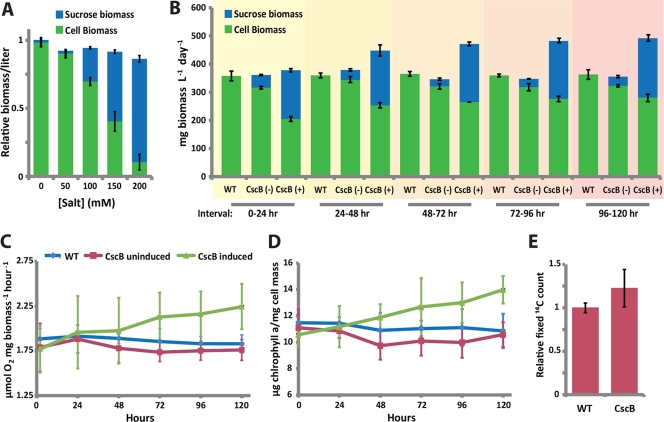
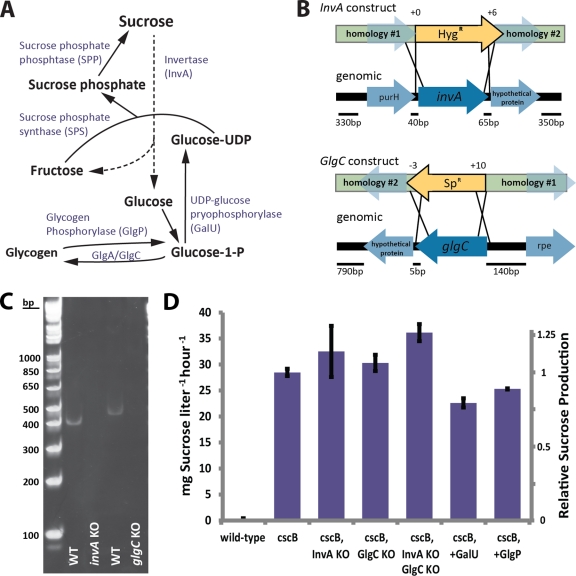
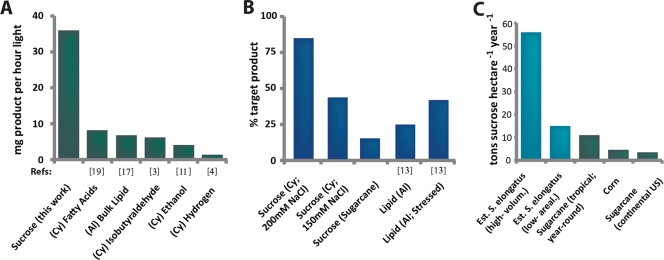
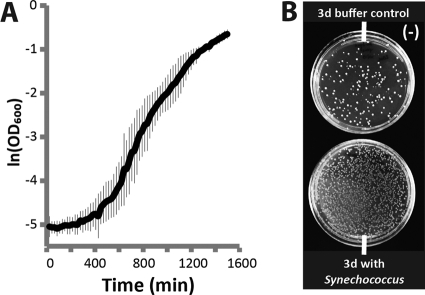
The bioindustrial production of fuels, chemicals, and therapeutics typically relies upon carbohydrate inputs derived from agricultural plants, resulting in the entanglement of food and chemical commodity markets. We demonstrate the efficient production of sucrose from a cyanobacterial species, Synechococcus elongatus, heterologously expressing a symporter of protons and sucrose (cscB). cscB-expressing cyanobacteria export sucrose irreversibly to concentrations of >10 mM without culture toxicity. Moreover, sucrose-exporting cyanobacteria exhibit increased biomass production rates relative to wild-type strains, accompanied by enhanced photosystem II activity, carbon fixation, and chlorophyll content. The genetic modification of sucrose biosynthesis pathways to minimize competing glucose- or sucrose-consuming reactions can further improve sucrose production, allowing the export of sucrose at rates of up to 36.1 mg liter(-1) h illumination(-1). This rate of production exceeds that of previous reports of targeted, photobiological production from microbes. Engineered S. elongatus produces sucrose in sufficient quantities (up to ∼80% of total biomass) such that it may be a viable alternative to sugar synthesis from terrestrial plants, including sugarcane.
Figures
References
-
- Abramson J, Iwata S, Kaback HR. 2004. Lactose permease as a paradigm for membrane transport proteins (review). Mol. Membr. Biol. 21:227–236 - PubMed
-
- Anonymous 2011, posting date World food situation. Food and Agriculture Organization of the United Nations http://www.fao.org/worldfoodsituation/wfs-home/en/
-
- Atsumi S, Higashide W, Liao JC. 2009. Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nat. Biotechnol. 27:1177–1180 - PubMed
-
- Bandyopadhyay A, Stockel J, Min H, Sherman LA, Pakrasi HB. 2010. High rates of photobiological H2 production by a cyanobacterium under aerobic conditions. Nat. Commun. 1:139. - PubMed
-
- Barford D, Hu SH, Johnson LN. 1991. Structural mechanism for glycogen phosphorylase control by phosphorylation and AMP. J. Mol. Biol. 218:233–260 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
