

Experimental evolution of multicellularity
- PMID: 22307617
- PMCID: PMC3277146
- DOI: 10.1073/pnas.1115323109
Experimental evolution of multicellularity
Abstract
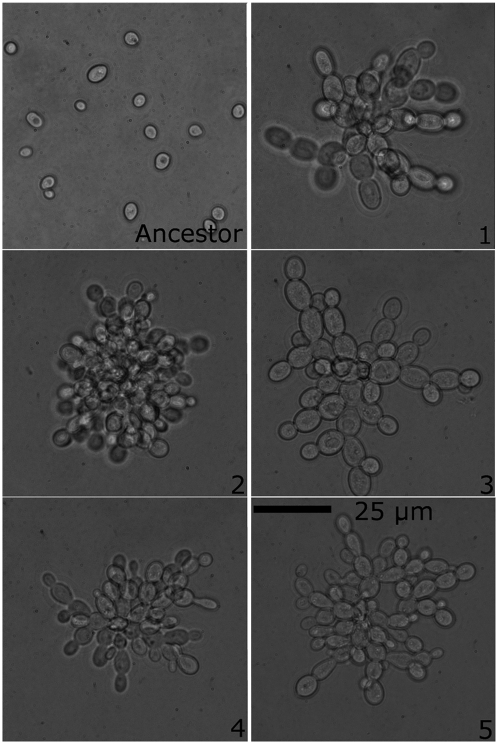
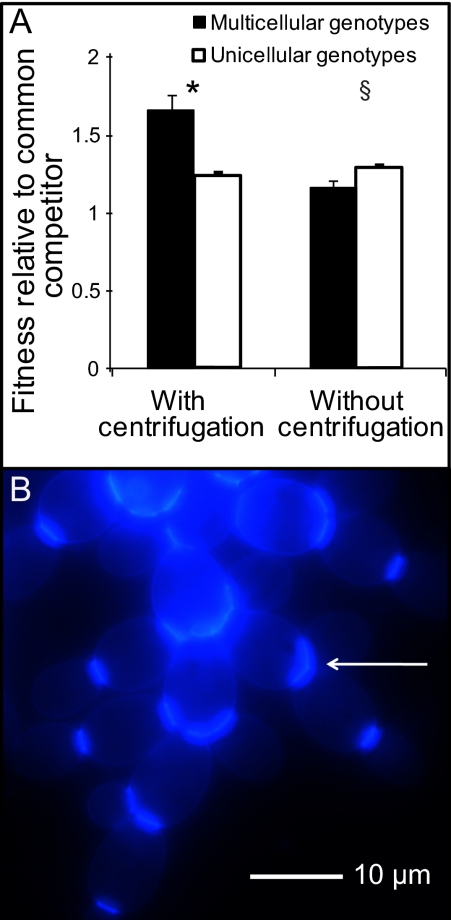
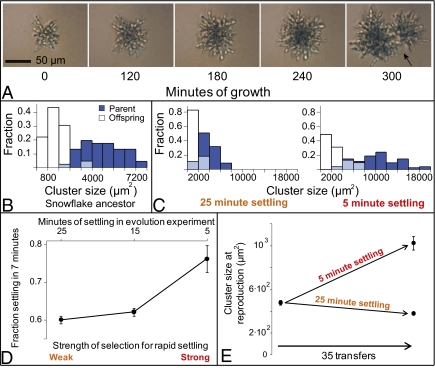
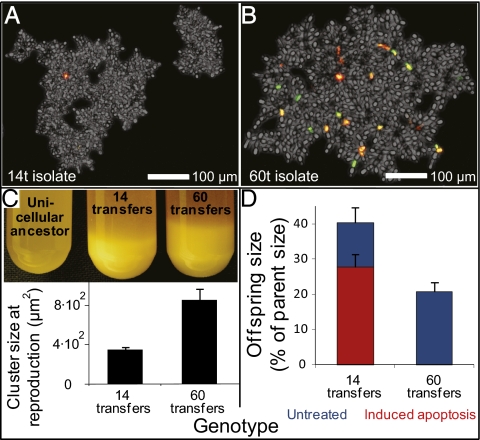
Multicellularity was one of the most significant innovations in the history of life, but its initial evolution remains poorly understood. Using experimental evolution, we show that key steps in this transition could have occurred quickly. We subjected the unicellular yeast Saccharomyces cerevisiae to an environment in which we expected multicellularity to be adaptive. We observed the rapid evolution of clustering genotypes that display a novel multicellular life history characterized by reproduction via multicellular propagules, a juvenile phase, and determinate growth. The multicellular clusters are uniclonal, minimizing within-cluster genetic conflicts of interest. Simple among-cell division of labor rapidly evolved. Early multicellular strains were composed of physiologically similar cells, but these subsequently evolved higher rates of programmed cell death (apoptosis), an adaptation that increases propagule production. These results show that key aspects of multicellular complexity, a subject of central importance to biology, can readily evolve from unicellular eukaryotes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
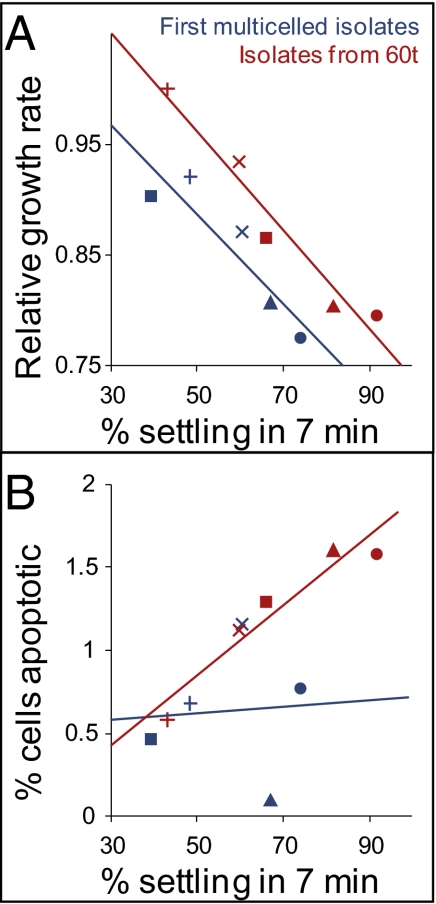
 , replicate population 7; ●, replicate population 8;
, replicate population 7; ●, replicate population 8;  replicate population 9). (B) The frequency of apoptotic cells (measured by dihydrorhodamine 123 staining for reactive oxygen species) was not correlated with settling rate in the first snowflake genotypes to evolve in each population (r2 = 0.005). By 60 transfers, however, settling rate and apoptosis are highly correlated (r2 = 0.91).
replicate population 9). (B) The frequency of apoptotic cells (measured by dihydrorhodamine 123 staining for reactive oxygen species) was not correlated with settling rate in the first snowflake genotypes to evolve in each population (r2 = 0.005). By 60 transfers, however, settling rate and apoptosis are highly correlated (r2 = 0.91).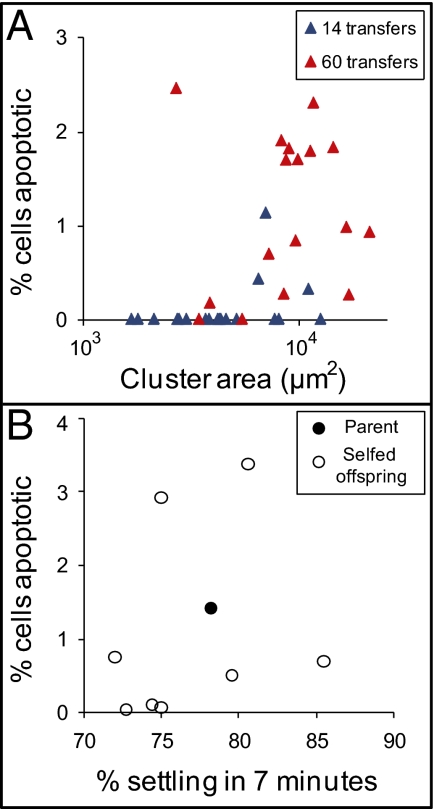
References
-
- Maynard Smith J, Szathmáry E. The Major Transitions in Evolution. New York: Oxford University Press; 1995.
-
- Kirk DL. A twelve-step program for evolving multicellularity and a division of labor. Bioessays. 2005;27:299–310. - PubMed
-
- Merlo LMF, Pepper JW, Reid BJ, Maley CC. Cancer as an evolutionary and ecological process. Nat Rev Cancer. 2006;6:924–935. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
