

Toll-like receptor 5 (TLR5), IL-1β secretion, and asparagine endopeptidase are critical factors for alveolar macrophage phagocytosis and bacterial killing
- PMID: 22307620
- PMCID: PMC3277124
- DOI: 10.1073/pnas.1108464109
Toll-like receptor 5 (TLR5), IL-1β secretion, and asparagine endopeptidase are critical factors for alveolar macrophage phagocytosis and bacterial killing
Abstract
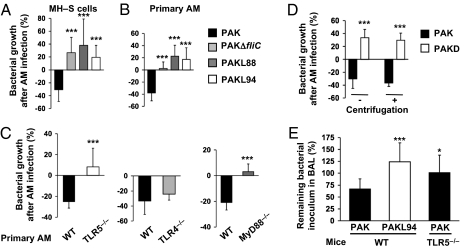
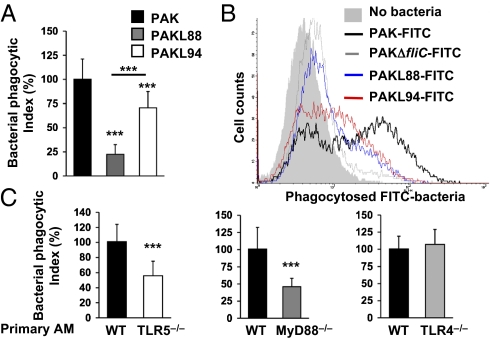
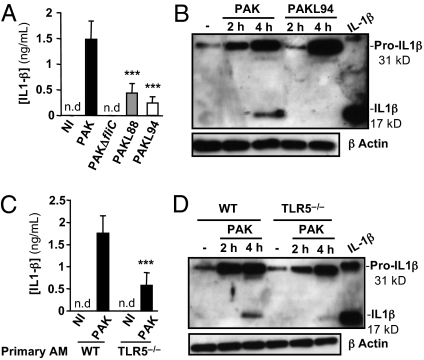
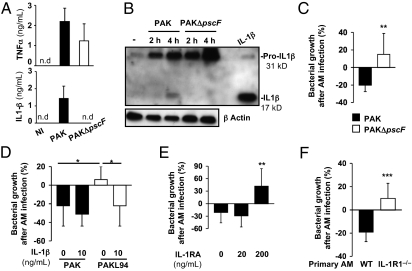
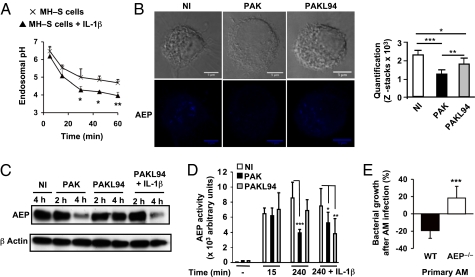
A deficit in early clearance of Pseudomonas aeruginosa (P. aeruginosa) is crucial in nosocomial pneumonia and in chronic lung infections. Few studies have addressed the role of Toll-like receptors (TLRs), which are early pathogen associated molecular pattern receptors, in pathogen uptake and clearance by alveolar macrophages (AMs). Here, we report that TLR5 engagement is crucial for bacterial clearance by AMs in vitro and in vivo because unflagellated P. aeruginosa or different mutants defective in TLR5 activation were resistant to AM phagocytosis and killing. In addition, the clearance of PAK (a wild-type P. aeruginosa strain) by primary AMs was causally associated with increased IL-1β release, which was dramatically reduced with PAK mutants or in WT PAK-infected primary TLR5(-/-) AMs, demonstrating the dependence of IL-1β production on TLR5. We showed that this IL-1β production was important in endosomal pH acidification and in inducing the killing of bacteria by AMs through asparagine endopeptidase (AEP), a key endosomal cysteine protease. In agreement, AMs from IL-1R1(-/-) and AEP(-/-) mice were unable to kill P. aeruginosa. Altogether, these findings demonstrate that TLR5 engagement plays a major role in P. aeruginosa internalization and in triggering IL-1β formation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Mogayzel PJ, Jr, Flume PA. Update in cystic fibrosis 2009. Am J Respir Crit Care Med. 2010;181:539–544. - PubMed
-
- Gordon S. The macrophage: Past, present and future. Eur J Immunol. 2007;37(Suppl 1):S9–S17. - PubMed
-
- Di A, et al. CFTR regulates phagosome acidification in macrophages and alters bactericidal activity. Nat Cell Biol. 2006;8:933–944. - PubMed
-
- Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat Immunol. 2010;11:373–384. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
