

Revisiting the ionic mechanisms of early afterdepolarizations in cardiomyocytes: predominant by Ca waves or Ca currents?
- PMID: 22307670
- PMCID: PMC3330805
- DOI: 10.1152/ajpheart.00742.2011
Revisiting the ionic mechanisms of early afterdepolarizations in cardiomyocytes: predominant by Ca waves or Ca currents?
Abstract
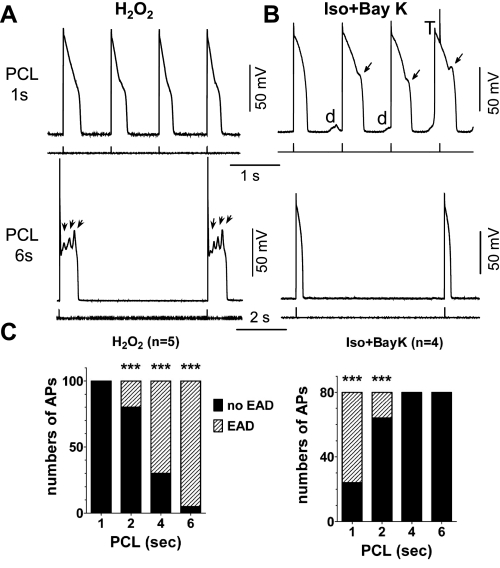
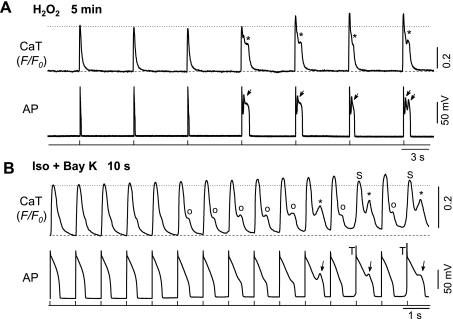
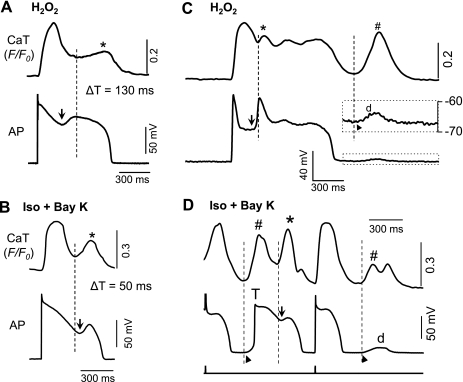
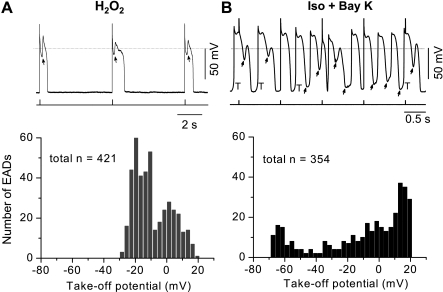
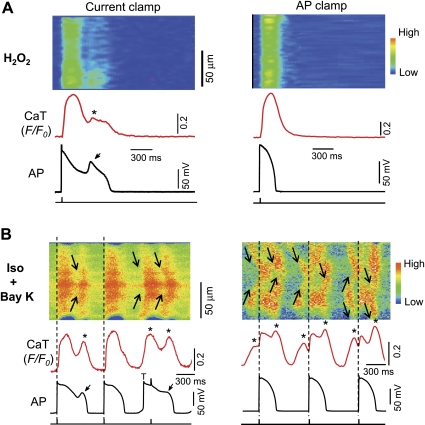
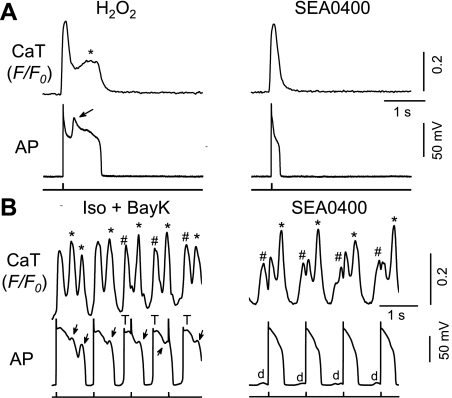
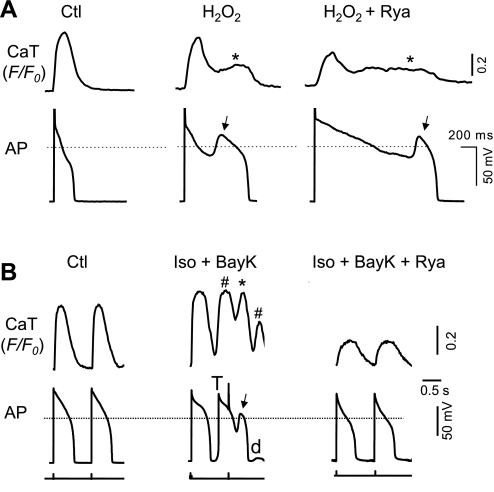
Early afterdepolarizations (EADs) have been implicated in severe cardiac arrhythmias and sudden cardiac deaths. However, the mechanism(s) for EAD genesis, especially regarding the relative contribution of Ca(2+) wave (CaW) vs. L-type Ca current (I(Ca,L)), still remains controversial. In the present study, we simultaneously recorded action potentials (APs) and intracellular Ca(2+) images in isolated rabbit ventricular myocytes and systematically compared the properties of EADs in the following two pharmacological models: 1) hydrogen peroxide (H(2)O(2); 200 μM); and 2) isoproterenol (100 nM) and BayK 8644 (50 nM) (Iso + BayK). We assessed the rate dependency of EADs, the temporal relationship between EADs and corresponding CaWs, the distribution of EADs over voltage, and the effects of blockers of I(Ca,L), Na/Ca exchangers, and ryanodine receptors. The most convincing evidence came from the AP-clamp experiment, in which the cell membrane clamp was switched from current clamp to voltage clamp using a normal AP waveform without EAD; CaWs disappeared in the H(2)O(2) model, but persisted in the Iso + BayK model. We postulate that, although CaWs and reactivation of I(Ca,L) may act synergistically in either case, reactivation of I(Ca,L) plays a predominant role in EAD genesis under oxidative stress (H(2)O(2) model), while spontaneous CaWs are a predominant cause for EADs under Ca(2+) overload condition (Iso + BayK model).
Figures
References
-
- Bechem M, Hoffmann H. The molecular mode of action of the Ca agonist (−)BAY K 8644 on the cardiac Ca channel. Pflügers Arch 424: 343–353, 1993 - PubMed
-
- Binah O, Rosen MR. Mechanisms of ventricular arrhythmias. Circulation 85: I25–31, 1992 - PubMed
-
- Birinyi P, Acsai K, Banyasz T, Toth A, Horvath B, Virag L, Szentandrassy N, Magyar J, Varro A, Fulop F, Nanasi PP. Effects of SEA0400 and KB-R7943 on Na+/Ca2+ exchange current and L-type Ca2+ current in canine ventricular cardiomyocytes. Naunyn Schmiedebergs Arch Pharmacol 372: 63–70, 2005 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous