

Identification of mechanism that couples multisite phosphorylation of Yes-associated protein (YAP) with transcriptional coactivation and regulation of apoptosis
- PMID: 22308035
- PMCID: PMC3308740
- DOI: 10.1074/jbc.M111.296954
Identification of mechanism that couples multisite phosphorylation of Yes-associated protein (YAP) with transcriptional coactivation and regulation of apoptosis
Erratum in
-
Identification of a mechanism that couples multisite phosphorylation of Yes-associated protein (YAP) with transcriptional coactivation and regulation of apoptosis.J Biol Chem. 2016 Feb 26;291(9):4844-5. doi: 10.1074/jbc.A111.296954. J Biol Chem. 2016. PMID: 26921413 Free PMC article. No abstract available.
Abstract
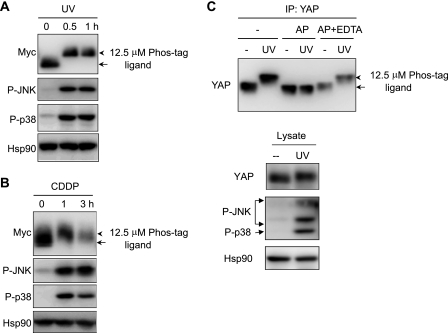
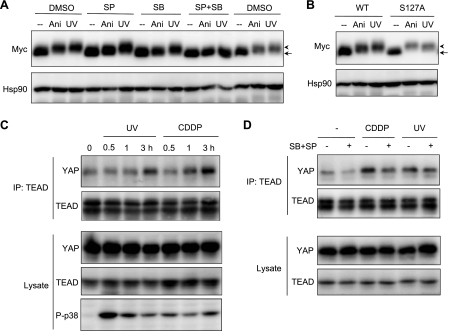
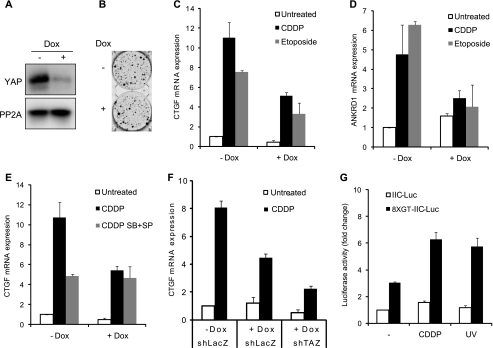
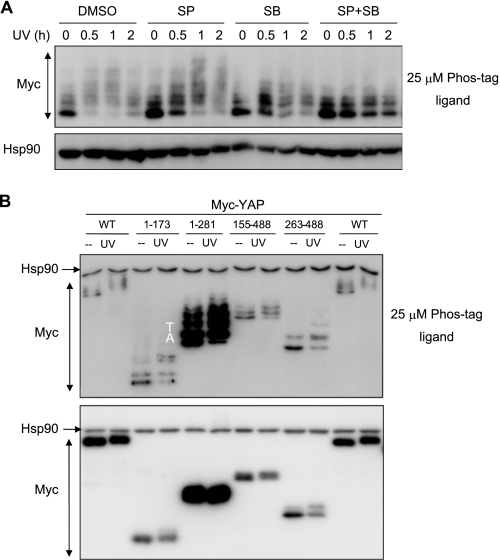
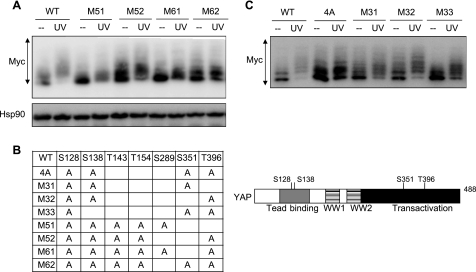
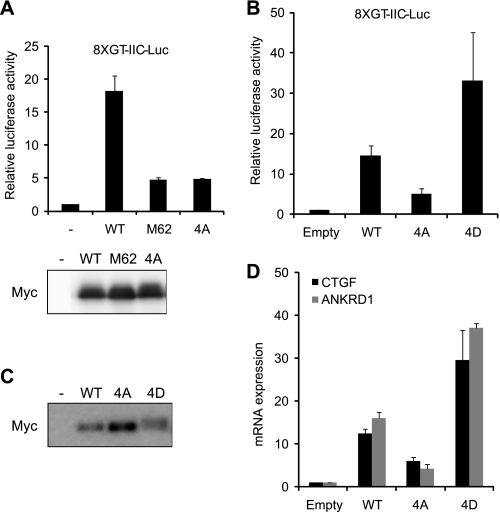
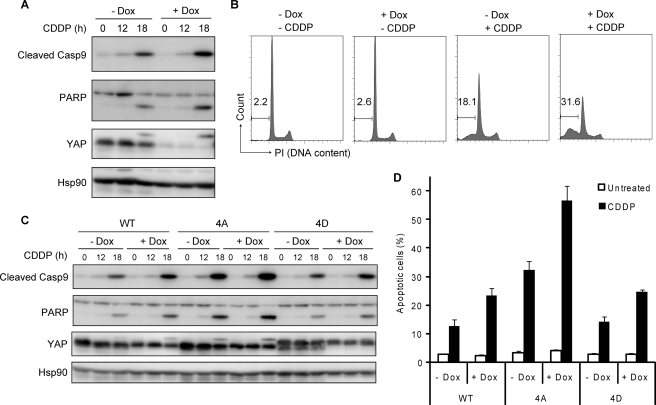
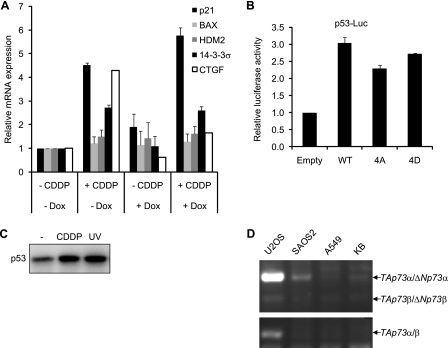
The transcriptional coactivator Yes-associated protein (YAP) has been implicated in tumorigenesis by regulating cell proliferation and apoptosis. YAP interacts with the transcription factor TEAD and is essential in mediating TEAD-dependent gene expression. Here we show that YAP is hyperphosphorylated and activated in response to genotoxic stress such as UV irradiation and cisplatin treatment. Using high resolution mobility shift assay for phosphorylated proteins, we identified multiple sites of phosphorylation induced by UV irradiation. Pretreatment with p38 and JNK inhibitors completely suppressed the mobility retardation of phosphorylated YAP in UV-irradiated cells. Co-immunoprecipitation experiments showed that the physical interaction of YAP with TEAD was markedly enhanced by UV irradiation or CDDP treatment but suppressed by pretreatment with p38 and JNK inhibitors. Similarly, pretreatment with p38 and JNK inhibitors suppressed the expression of YAP/TEAD target genes, which were elevated on exposure to genotoxic stress. Using phosphomimetic and phosphorylation-deficient YAP mutants, we showed that the coactivator activity of YAP correlated with its state of phosphorylation and sensitivity to cisplatin-induced apoptosis. Our results demonstrate that multisite phosphorylation of YAP induces YAP/TEAD-dependent gene expression and provides a mechanism by which YAP regulates apoptosis differently depending on cellular context.
Figures
References
-
- Conlon I., Raff M. (1999) Size control in animal development. Cell 96, 235–244 - PubMed
-
- McClatchey A. I., Giovannini M. (2005) Membrane organization and tumorigenesis. The NF2 tumor suppressor, Merlin. Genes Dev. 19, 2265–2277 - PubMed
-
- St John M. A., Tao W., Fei X., Fukumoto R., Carcangiu M. L., Brownstein D. G., Parlow A. F., McGrath J., Xu T. (1999) Mice deficient of Lats1 develop soft-tissue sarcomas, ovarian tumors, and pituitary dysfunction. Nat. Genet. 21, 182–186 - PubMed
-
- Zhou D., Conrad C., Xia F., Park J. S., Payer B., Yin Y., Lauwers G. Y., Thasler W., Lee J. T., Avruch J., Bardeesy N. (2009) Mst1 and Mst2 maintain hepatocyte quiescence and suppress hepatocellular carcinoma development through inactivation of the Yap1 oncogene. Cancer Cell 16, 425–438 - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
