

RNA virus microRNA that mimics a B-cell oncomiR
- PMID: 22308400
- PMCID: PMC3286953
- DOI: 10.1073/pnas.1116107109
RNA virus microRNA that mimics a B-cell oncomiR
Abstract
MicroRNAs (miRNAs) are small RNAs that play a regulatory role in numerous and diverse eukaryotic cellular processes. Virus-encoded miRNAs have garnered much interest, although the functions of most remain to be deciphered. To date, readily detectable, evolutionarily conserved natural miRNAs have only been identified from viruses with DNA genomes. Combined with the fact that most miRNAs are generated from endonucleolytic cleavage of longer transcripts, this finding has led to a common conception that naturally occurring RNA viruses will not encode miRNAs to avoid unproductive cleavage of their genomes or mRNAs. Here we demonstrate that the bovine leukemia virus (BLV), a retrovirus with an RNA genome, encodes a conserved cluster of miRNAs that are transcribed by RNA polymerase III (pol III). Thus, the BLV miRNAs avoid the conundrum of genome/mRNA cleavage because only the subgenomic pol III transcripts are efficiently processed into miRNAs. BLV infection is strongly associated with B-cell tumors in cattle. Because most cells in BLV-associated tumors express little viral mRNAs or proteins, exactly how BLV contributes to tumorigenesis has remained a decades-long unsolved mystery. One BLV miRNA, BLV-miR-B4, shares partial sequence identity and shared common targets with the host miRNA, miR-29. As miR-29 overexpression is associated with B-cell neoplasms that resemble BLV-associated tumors, our findings suggest a possible mechanism contributing to BLV-induced tumorigenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
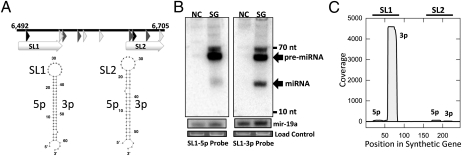
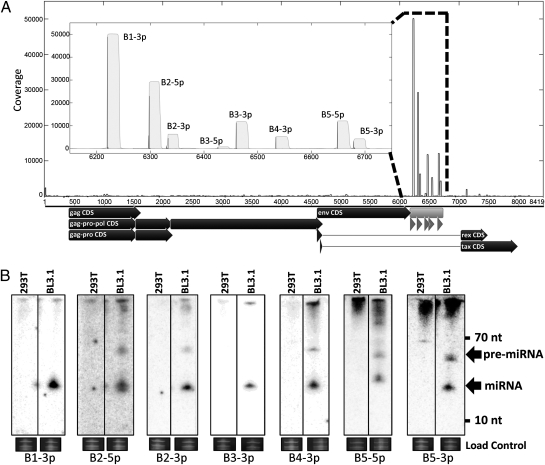
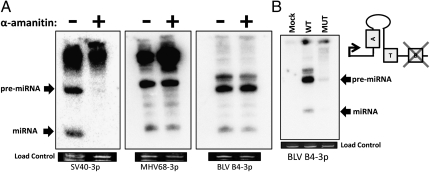
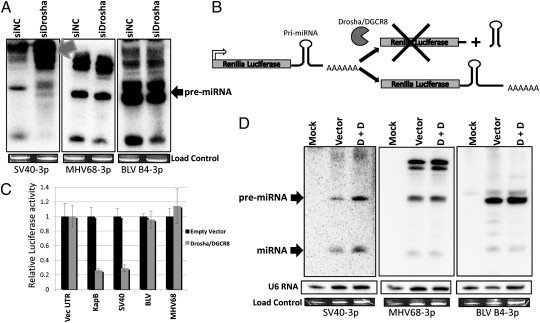
Comment in
-
MicroRNA expression by an oncogenic retrovirus.Proc Natl Acad Sci U S A. 2012 Feb 21;109(8):2695-6. doi: 10.1073/pnas.1200328109. Epub 2012 Feb 2. Proc Natl Acad Sci U S A. 2012. PMID: 22308505 Free PMC article. No abstract available.
-
Retrovirus infected cells contain viral microRNAs.Retrovirology. 2013 Feb 7;10:15. doi: 10.1186/1742-4690-10-15. Retrovirology. 2013. PMID: 23391025 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
