

Phospholipase C-related but catalytically inactive protein (PRIP) modulates synaptosomal-associated protein 25 (SNAP-25) phosphorylation and exocytosis
- PMID: 22311984
- PMCID: PMC3322965
- DOI: 10.1074/jbc.M111.294645
Phospholipase C-related but catalytically inactive protein (PRIP) modulates synaptosomal-associated protein 25 (SNAP-25) phosphorylation and exocytosis
Abstract
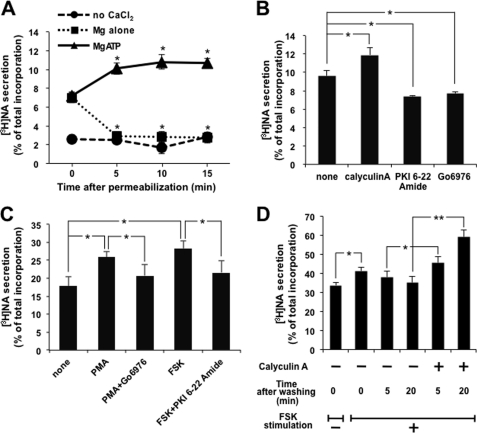
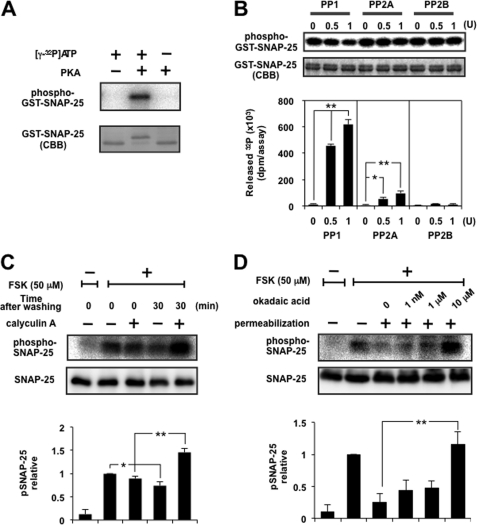
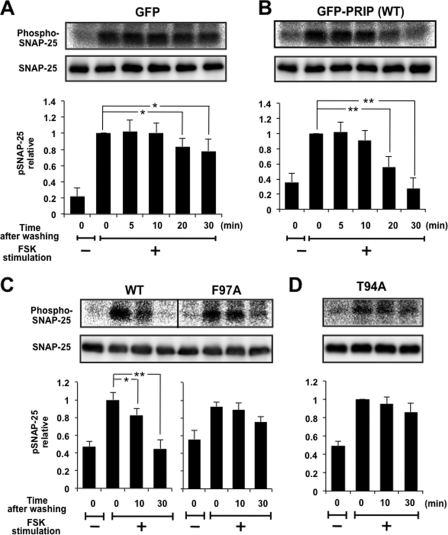
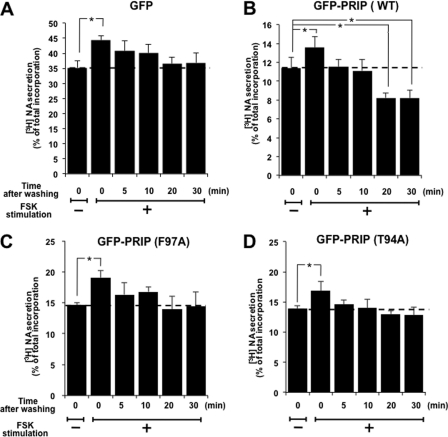
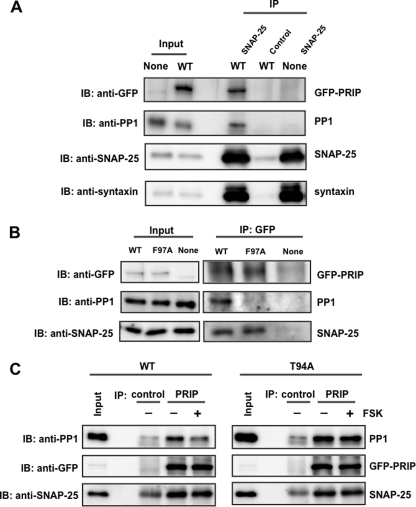
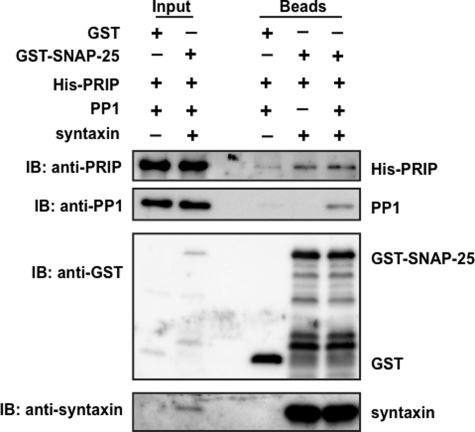
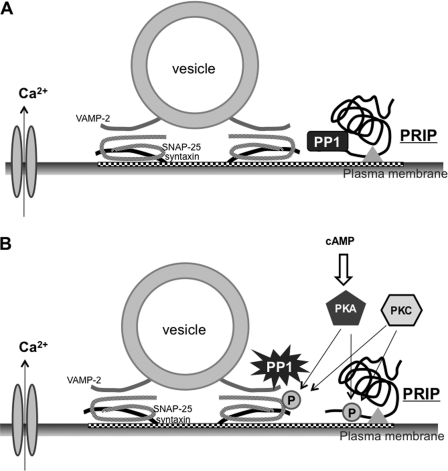
Exocytosis is one of the most fundamental cellular events. The basic mechanism of the final step, membrane fusion, is mediated by the formation of the SNARE complex, which is modulated by the phosphorylation of proteins controlled by the concerted actions of protein kinases and phosphatases. We have previously shown that a protein phosphatase-1 (PP1) anchoring protein, phospholipase C-related but catalytically inactive protein (PRIP), has an inhibitory role in regulated exocytosis. The current study investigated the involvement of PRIP in the phospho-dependent modulation of exocytosis. Dephosphorylation of synaptosome-associated protein of 25 kDa (SNAP-25) was mainly catalyzed by PP1, and the process was modulated by wild-type PRIP but not by the mutant (F97A) lacking PP1 binding ability in in vitro studies. We then examined the role of PRIP in phospho-dependent regulation of exocytosis in cell-based studies using pheochromocytoma cell line PC12 cells, which secrete noradrenalin. Exogenous expression of PRIP accelerated the dephosphorylation process of phosphorylated SNAP-25 after forskolin or phorbol ester treatment of the cells. The phospho-states of SNAP-25 were correlated with noradrenalin secretion, which was enhanced by forskolin or phorbol ester treatment and modulated by PRIP expression in PC12 cells. Both SNAP-25 and PP1 were co-precipitated in anti-PRIP immunocomplex isolated from PC12 cells expressing PRIP. Collectively, together with our previous observation regarding the roles of PRIP in PP1 regulation, these results suggest that PRIP is involved in the regulation of the phospho-states of SNAP-25 by modulating the activity of PP1, thus regulating exocytosis.
Figures

References
-
- Banerjee A., Barry V. A., DasGupta B. R., Martin T. F. (1996) N-Ethylmaleimide-sensitive factor acts at a prefusion ATP-dependent step in Ca2+-activated exocytosis. J. Biol. Chem. 271, 20223–20226 - PubMed
-
- Sutton R. B., Fasshauer D., Jahn R., Brunger A. T. (1998) Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 Å resolution. Nature 395, 347–353 - PubMed
-
- Weber T., Zemelman B. V., McNew J. A., Westermann B., Gmachl M., Parlati F., Söllner T. H., Rothman J. E. (1998) SNAREpins. Minimal machinery for membrane fusion. Cell 92, 759–772 - PubMed
-
- Jahn R., Lang T., Südhof T. C. (2003) Membrane fusion. Cell 112, 519–533 - PubMed
-
- Söllner T. H. (2003) Regulated exocytosis and SNARE function (Review). Mol. Membr. Biol. 20, 209–220 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
