

Inhibition of pyrimidine synthesis reverses viral virulence factor-mediated block of mRNA nuclear export
- PMID: 22312003
- PMCID: PMC3275370
- DOI: 10.1083/jcb.201107058
Inhibition of pyrimidine synthesis reverses viral virulence factor-mediated block of mRNA nuclear export
Abstract
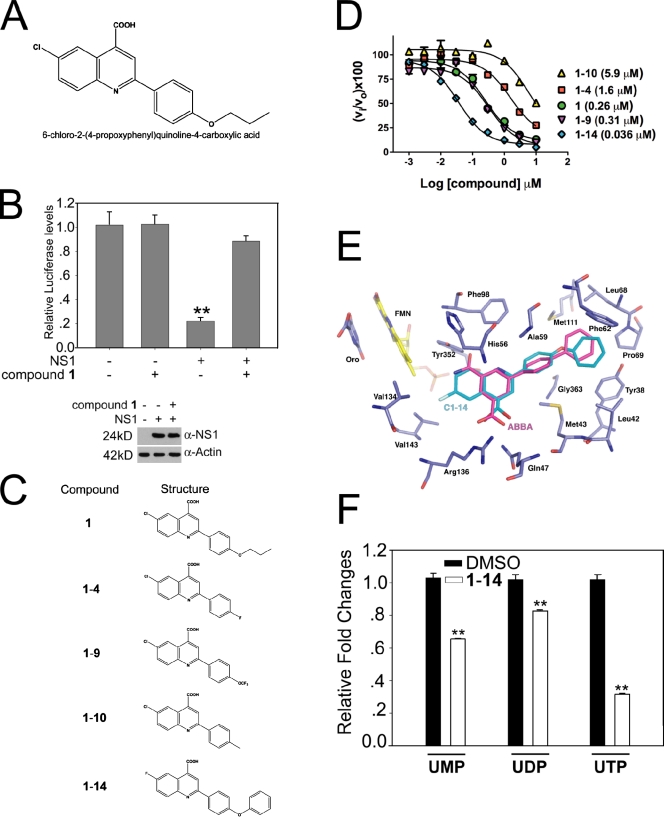
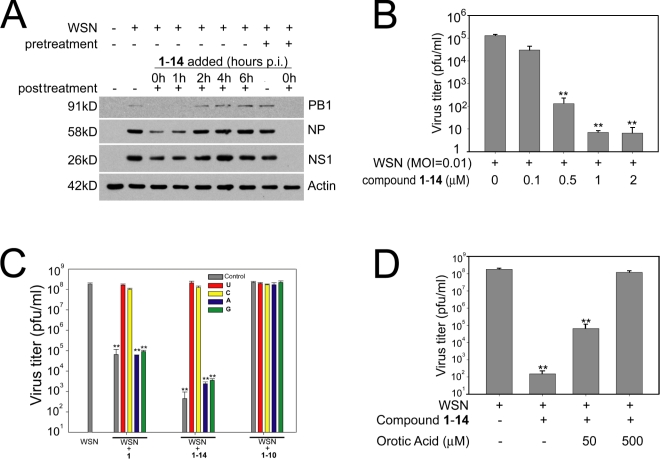
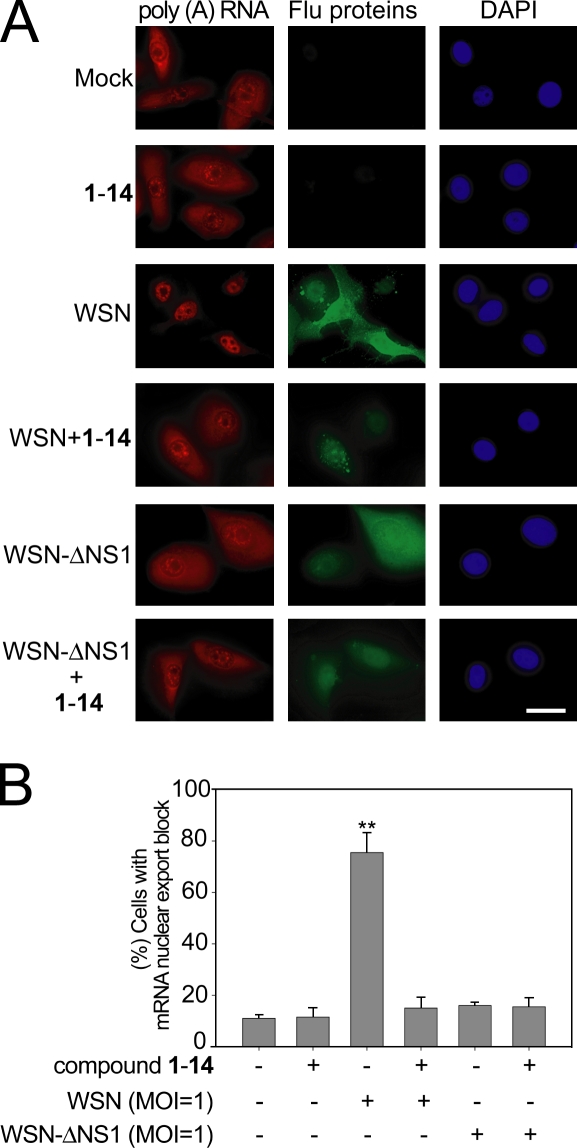
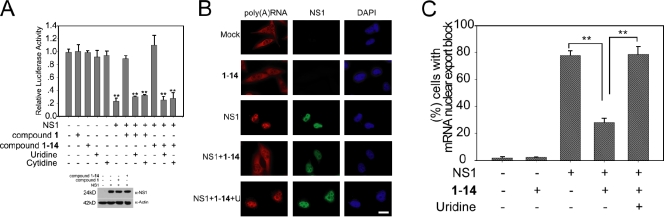
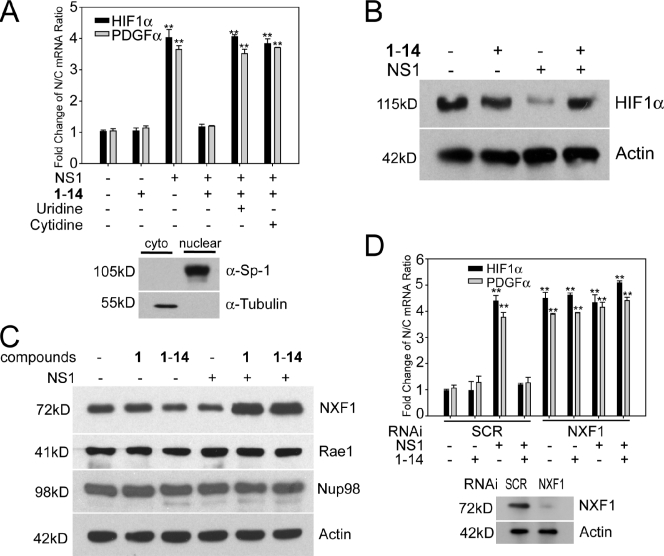
The NS1 protein of influenza virus is a major virulence factor essential for virus replication, as it redirects the host cell to promote viral protein expression. NS1 inhibits cellular messenger ribonucleic acid (mRNA) processing and export, down-regulating host gene expression and enhancing viral gene expression. We report in this paper the identification of a nontoxic quinoline carboxylic acid that reverts the inhibition of mRNA nuclear export by NS1, in the absence or presence of the virus. This quinoline carboxylic acid directly inhibited dihydroorotate dehydrogenase (DHODH), a host enzyme required for de novo pyrimidine biosynthesis, and partially reduced pyrimidine levels. This effect induced NXF1 expression, which promoted mRNA nuclear export in the presence of NS1. The release of NS1-mediated mRNA export block by DHODH inhibition also occurred in the presence of vesicular stomatitis virus M (matrix) protein, another viral inhibitor of mRNA export. This reversal of mRNA export block allowed expression of antiviral factors. Thus, pyrimidines play a necessary role in the inhibition of mRNA nuclear export by virulence factors.
© 2012 Zhang et al.
Figures
References
-
- Batt D.G., Copeland N.G., Dowling R.L., Gardner T.L., Jones E.A., Orwat M.J., Pinto D.J., Pitts W.J., Magolda R.L., Jaffee B.D. 1995. Immunosuppressive structure-activity relationships of Brequinar and related cinchoninic acid derivatives. Bioorg. Med. Chem. Lett. 5:1549–1554 10.1016/0960-894X(95)00252-O - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- C06 RR015437/RR/NCRR NIH HHS/United States
- R01 AI089539/AI/NIAID NIH HHS/United States
- R01 AI046954/AI/NIAID NIH HHS/United States
- R01 GM094314/GM/NIGMS NIH HHS/United States
- R01 GM07159/GM/NIGMS NIH HHS/United States
- U19 AI083025/AI/NIAID NIH HHS/United States
- P30 CA142543/CA/NCI NIH HHS/United States
- U54 AI057158/AI/NIAID NIH HHS/United States
- U19AI083025/AI/NIAID NIH HHS/United States
- R01 AI079110/AI/NIAID NIH HHS/United States
- 2R56AI53680/AI/NIAID NIH HHS/United States
- P01AI058113/AI/NIAID NIH HHS/United States
- R01AI28900/AI/NIAID NIH HHS/United States
- U54AI057158/AI/NIAID NIH HHS/United States
- R56 AI053680/AI/NIAID NIH HHS/United States
- R01AI046954/AI/NIAID NIH HHS/United States
- R01 AI028900/AI/NIAID NIH HHS/United States
- C06-RR15437/RR/NCRR NIH HHS/United States
- R01AI089539/AI/NIAID NIH HHS/United States
- HHSN266200700010C/AI/NIAID NIH HHS/United States
- P01 AI058113/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
